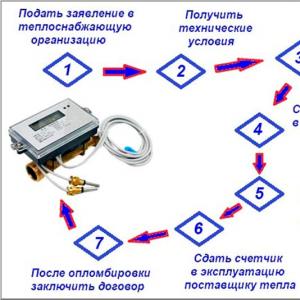
Selkärankaisten elinjärjestelmät. Maan historia Selkärankainen järjestelmä
Selkärankaisten esi-isät, ilmeisesti olivat lähellä primitiivisiä pohjaeläin-pelagisten ei-kalloeläinten muotoja, jotka eivät olleet vielä hankkineet eteisonteloa. Oletettavasti nämä olivat pieniä vesieläimiä, joilla oli tyypillisiä sointujen merkkejä, mutta niillä ei vielä ollut vahvaa sisäistä luurankoa ja voimakkaita lihaksia. Ne luultavasti erosivat alkuperäisistä ryhmistä ordovikiassa, eli ainakin 500 miljoonaa vuotta sitten. Emme kuitenkaan tunne niitä, eivätkä niitä todennäköisesti koskaan löydetä, koska niiden pieni koko ja kiinteiden luustomuodostelmien puuttuminen tekevät fossiilisten jäänteiden säilymisen epätodennäköiseksi.
Huonosti säilyneitä primitiivisten, mutta epäilemättä jo täysin kehittyneiden selkärankaisten (scutellous agnathans) jäänteitä tunnetaan Ordovikian-Alasilurian esiintymistä (noin 450 miljoonaa vuotta vanhoja). He asuivat luultavasti makeissa vesistöissä, ja niiden jäännökset kuljetettiin virtauksen mukana matalille merenlahdille, jonne ne sijoitettiin. Niiden olemassaoloa jokien suistoissa ja meren suolattomilla alueilla ei ole poissuljettu. On erittäin mielenkiintoista, että selkärankaisten jäänteitä alkaa esiintyä tyypillisissä meriesiintymissä vasta devonikauden puolivälistä lähtien, eli noin 350 ml:n ikäisissä esiintymissä. vuosia sitten ja myöhemmin. Nämä paleontologiset tiedot viittaavat siihen, että selkärankaisten muodostuminen ei tapahtunut merissä, vaan makeissa vesissä. Mikä voisi olla syy merellisten sointujen - selkärankaisten esi-isimpien - siirtymiseen meristä makeaan veteen?
punasilmäinen puusammakko(Agalychnis callidryas)
Paleontologisten tietojen perusteella Ordovician ja Silurian aikana merten ja valtamerten pohjabiokenoosit olivat runsaasti erilaisia eläimiä - matoja, nilviäisiä, äyriäisiä ja piikkinahkaisia. Pohjassa oli luultavasti lukuisia alempia sointuja, mukaan lukien vaippakappaleet; Täällä asui myös sellaiset voimakkaat saalistajat, kuten suuret pääjalkaiset ja jättiläisäyriäisskorpionit (pituudeltaan 3-5 m). Uuden suuren ryhmän (selkärankaisten alatyyppi) ilmaantuminen tällaisten kyllästyneiden biokenoosien olosuhteissa, joissa on kovaa kilpailua, vaikuttaa epätodennäköiseltä. Samaan aikaan makeissa vesistöissä oli jo melko rikas kasvisto (pääasiassa levät) ja huomattava määrä erilaisia selkärangattomia; suuria ja vahvoja saalistajia oli vähän. Siksi selkärankaisten esi-isien tunkeutuminen tänne ja niiden myöhempi kehitys näyttää mahdolliselta. Meriin verrattuna makeissa vesissä oli kuitenkin myös epäsuotuisia ominaisuuksia, jotka estivät meren eliöiden kulkeutumisen. Merkittävä määrä veteen liuenneita suoloja uhkasi meren hyökkääjiä vettä läpäisevillä peitteillä ruumiin liiallisella kastelulla, suolakoostumuksen jyrkällä rikkomisella ja kudosten osmoottisella paineella, minkä olisi pitänyt johtaa yleiseen aineenvaihduntahäiriöön. Makeavesistöille on mereen verrattuna ominaista epävakaampi kemia (mukaan lukien jyrkät happipitoisuuden vaihtelut) ja vaihteleva lämpötilajärjestelmä. Jokiin asettuminen vaati myös suurta liikkuvuutta pysyäkseen tietyllä alueella ja vastustaakseen virran ajautumista.
Mutta petoeläinten heikko paine, ravinnon suhteellinen runsaus ja vähemmän akuutti kilpailu makean veden biokenoosissa loivat mahdollisuudet joidenkin sointujen edustajien tunkeutumiseen ja niiden johdonmukaiseen sopeutumiseen uusiin olosuhteisiin. Ilmeisesti tällä tavalla tapahtui selkärankaisten muodostuminen, joiden esi-isät tunkeutuivat ensin suistoon, siirtyivät sitten jokien alajuoksulle ja alkoivat nousta ylävirtaan, asuttaen järviä. Tällaisissa paikoissa olisi voinut syntyä selkärankaisille tyypillisiä munuaisia, jotka pystyivät suodattamaan valtavia määriä vettä kehosta, eliminoimaan kehon mehujen osmoottisen paineen rikkoutumisen ja estämään uusissa olosuhteissa puutteellisten suolojen häviämisen. Makean vesistön happipitoisuuden kausivaihtelut vaativat hengityselinten parantamista, jonka evoluutioreaktio oli ilmeisesti kidusten ilmaantuminen. Tarve voittaa virtojen voima paransi liikkuvuutta: kehittyi vahvempi, mutta joustavuutta säilyttävä aksiaalinen luuranko selkärangan muodossa - tuki motoristen lihasten kasvavalle massalle. Suurempi liikkuvuus puolestaan vaati hermoston ja aistielinten, verenkierto-, ruoansulatus- ja eritysjärjestelmien komplikaatioita. Lihaslihaskompleksin vahvistumista helpotti kehon koon kasvu, mikä puolestaan mahdollisti liikkeen nopeuden lisäämisen, koska pitkän kehon harvinaisemmat värähtelyt ovat mekaanisesti hyödyllisiä kuin toistuvat lyhyen rungon värähtelyt. Liikkumisnopeuden lisääntyminen vedessä ja kehon absoluuttisten mittojen kasvu voidaan osoittaa kalojen esimerkillä.
Ensimmäiset selkärankaiset liittyvät leuattomiin ( Agnatha), ilmeisesti erotettu esi-isien ryhmistä suolattomissa suistoissa ja suurten jokien suussa. Tämä tapahtui Ordovikian lopussa - Silurian alussa. Heillä ei ollut leukoja ja parillisia eviä, he viettivät pohjaeläimen elämäntapaa, ruokkivat jätteitä ja ehkä pieniä, inaktiivisia pohjaeläimen selkärangattomia. Myöhemmin nämä eläimet saivat vahvan ulkokuoren, joka ilmeisesti suojeli niitä saalistavilta skorpionin kaltaisilta makean veden petoeläimiltä - eurypetridiltä ja auttoi samalla vähentämään makean veden tunkeutumista kehoon. Ordovikiassa - Silurialla (noin 400-450 miljoonaa vuotta sitten) syntyi ainakin kolme leuattomien scutellum-ryhmää, joista yhdestä alkoi ilmeisesti tuolloin erottua toinen selkärankaisten haara - leuallinen - Gnathostomata(panssaroitu kala - Placodermi). Toisin kuin leuattomien leuattomien esivanhempien, on todennäköistä, että he tunkeutuivat nopeammalla virtauksella jokiin, joissa valinta myötävaikutti vahvemman sisäisen luuston ja voimakkaiden lihasten muodostumiseen, parillisten evien muodostumiseen ja aistielinten parantamiseen. ja keskushermosto. Tämä lisäsi liikkuvuutta ja parempaa suuntautumista sekä mahdollisti asumisen ja erilaisten ruoka-aineiden metsästyksen nopeavirtaisissa vesissä. Vähitellen muodostui leukalaite - elin liikkuvan saaliin aktiiviseen sieppaamiseen.
Uskotaan, että primitiivinen selkärankainen ( Protocraniota) - leuattomien ja leuattomien esi-isä - omistettu kuin kalloton ( Acrania), jolla oli monimutkaisesti rei'itetty nielu, ja se oli suodatinta ruokkiva mikrofagi. Veden virtaus nielun läpi, jota tukivat segmentoimattomat kiduskaaret, kuten nyt elävä nahkiaisen toukka, hiekkamato, saatiin aikaan kidusonteloa puristavien lihasten supistumisella (uloshengitys); sisäänhengitys tapahtui seinien elastisen rentoutumisen aikana (kaaret); ruoka vangittiin limavirtauksella nielun alkuosassa. Seuraavassa evoluution vaiheessa kiduskaaret saivat nivelletyn rakenteen ja niihin liittyvät lihakset ilmestyivät laajentaen aktiivisesti kidusonteloa. Samaan aikaan, kun vesivirtaus lisääntyi, suuret ruokahiukkaset pystyivät pidättämään etukaaret. Lopuksi leuallisten selkärankaisten muodostumisen aikana yhden etummaisen haarakaaren (todennäköisesti kolmannen) alaosa (luultavasti kolmas) muutettiin alaleuaksi ja tämän kaaren yläosa (palatine-neliö) kiinnitettiin aksiaalinen kallo, joka muodostaa tartuntaelimen - leuat. Etukaaret ovat muuttuneet häpyrustoiksi (rustokaloilla on), kun taas takakaaret ovat säilyneet kiduslaitteiston tukena ( Lessertiseur, Robine, 1970).
Jo keski-devonilla (eli noin 320 miljoonaa vuotta sitten) sammakkoeläimet ( Amfibia). Hiilikaudella niitä edusti useita eri kokoisia, rakenteellisia ja ulkomuotoisia ryhmiä, jotka asettuivat makean veden rannikkoalueille, mutta sitten monet niistä katosivat; triasskaudella (noin 170-180 miljoonaa vuotta sitten) suuret sammakkoeläimet - stegokefalit - kuolivat sukupuuttoon.

Hiilikauden puolivälissä (noin 250-260 miljoonaa vuotta sitten) matelijat erosivat sammakkoeläimistä ( Reptilia). Koko mesozoisen aikakauden, yli 120 miljoonan vuoden ajan, he hallitsivat maapalloa ja hallitsivat menestyksekkäästi lähes kaikkia elämänalueita, mukaan lukien makeat ja merivedet ja ilma. Sopeutuessaan elämään erilaisissa elinympäristöissä matelijat jakautuivat 6-7 alaluokkaan. Matelijoiden kukinta edellytti muinaisten sammakkoeläinten sukupuuttoon. Liitukauden loppuun mennessä (noin 60-80 miljoonaa vuotta sitten) myös monet matelijaryhmät kuolivat sukupuuttoon suhteellisen nopeasti. Tähän päivään asti säilyneet lajit edustavat kolmen alaluokan suhteellisen köyhiä jäänteitä. Matelijoiden sukupuuttoon ei vaikuttanut ainoastaan haitalliset ilmastonmuutokset ja alppivuoristosykliä seurannut kasvillisuuden luonteen muutos, vaan myös lintujen ja nisäkkäiden intensiivinen lajiutuminen, joista oli tuolloin tullut vakavia kilpailijoita. mesozoisista matelijoista. Kuitenkin nämä kaksi korkeampien selkärankaisten luokkaa erosivat matelijoiden esi-isistään paljon aikaisemmin, pitkän aikaa (kymmeniä miljoonia vuosia) oli vain vähän ja ilmeisesti johti suhteellisen salaperäistä elämäntapaa.
Vasta liitukauden lopussa alkaa lintujen ja nisäkkäiden nopea kehitys ja samalla matelijoiden sukupuutto. Tätä edesauttoivat globaalit muutokset maapallolla (vuorenrakennusprosessit ja lisääntynyt tulivuoren aktiivisuus, mannerilmaston nousu monilla alueilla jne.; jopa oletuksia tehtiin kosmisten tekijöiden vaikutuksesta). Linnut ( Aves) erotettu erittäin järjestäytyneistä matelijoista - joistakin arkosaurusista - ilmeisesti triaskauden puolivälissä, vaikka vanhimmat ja alkeellisimmat linnut tunnetaan nykyään vain jurakauden (noin 135 miljoonaa vuotta vanhoista) esiintymistä. Joidenkin nykyaikaisten luokkien edustajia on jo löydetty myöhäisen liitukauden esiintymistä.
Nisäkkäät ( nisäkkäät) erottuivat yhdestä vanhimmista matelijaryhmistä - eläinmaisista matelijoista (alaluokka Theromorpha, seu synapsida); synapsidit syntyivät luultavasti hiilikauden puolivälissä, kokivat laajaa mukautuvaa säteilyä permikaudella ja katosivat triasskauden lopussa. Siksi nisäkkäiden muodostumisen olisi luultavasti pitänyt tapahtua triaskauden alussa (joidenkin paleontologien mukaan permikauden lopussa). Mesozoisten nisäkkäiden historia on huonosti tunnettu. Mesozoic-ajan (trias-jura) puolivälissä useat suhteellisen pian sukupuuttoon kuolleet ryhmät ilmeisesti jo erosivat (mutta niiden joukossa oli monotreemeja, jotka säilyivät meidän aikanamme). Pussaeläimet ja istukkaeläimet tunnetaan jurakaudesta, osa liitukaudesta. Istukan nisäkkäiden laajaa adaptiivista säteilyä ja nykyaikaisten luokkien muodostumista tapahtui jo kenozoisen aikakauden tertiaarikaudella (noin 60-40 miljoonaa vuotta sitten).
Tällaisesta vertailusta voidaan saada kuvaannollinen käsitys sointujen peräkkäisestä kehityksestä. Jos otamme yhden maapallon vuoden ajanjakson Maaplaneetan koko historian mittakaavaksi, niin elämän ilmaantuminen tapahtuu toukokuun lopussa - kesäkuun alussa, alempien selkärangattomien ilmaantuessa - klo. kesäkuun lopussa - heinäkuun alussa ja muun tyyppiset selkärangattomat ja alkeellisimmat sondit - syyskuun lopussa (paleotsoisen aikakauden kambrikausi). Lokakuun puolivälissä ilmestyvät ensimmäiset selkärankaiset - primitiiviset leuattomat (Ordovician loppu - Silurian alku) ja lokakuun lopussa (Silur) ensimmäiset leuattomat - primitiiviset kalat eristetään leuattomista. Ensimmäisen marraskuun lopussa - marraskuun toisen vuosikymmenen alussa (keski-devoni), ensimmäiset sammakkoeläimet erottuvat lohkoeväkaloista; luultavasti marraskuun viiden viimeisen päivän alussa (hiilikauden puolivälissä) ensimmäiset matelijat ilmestyvät ja marraskuun lopusta - joulukuun ensimmäisinä päivinä (permikausi) sammakkoeläinten sukupuutto ja matelijoiden kukinnan alku alkaa ja jatkuu joulukuun toisen vuosikymmenen loppuun (koko mesozoinen aikakausi). Triaskauden alussa (noin 3.–4. joulukuuta meidän mittakaavassa) muinaiset nisäkkäät erottuivat primitiivisistä matelijoista, ja saman ajanjakson lopussa (7.–8. joulukuuta) muinaiset linnut erosivat progressiivisista matelijoista - arkosaurusista.
Kuitenkin vasta joulukuun toisen vuosikymmenen lopussa (liitukauden lopussa) alkaa lintujen ja nisäkkäiden nopea kehitys ja monien mesotsoisten matelijaryhmien sukupuuttoon. Cenozoic aikakausi - korkeampien selkärankaisten nykyaikaisten ryhmien muodostumisaika - alkaa vasta joulukuun 23. päivän tienoilla ja monien nykyaikaisten perheiden muodostuminen - 28. joulukuuta (neogeenin alku). Pleistoseenin (kvaternaarin) alku (tässä mittakaavassa) osuu noin kello 18-20 31. joulukuuta; tämä on alkukantaisten (muinaisten) ihmislajien ja nykyaikaisten tai lähellä nykyaikaisten nisäkäs- ja lintulajien ilmaantumisen aika. Moderni mies ( Homo sapiens- järkevä mies) ilmestyi noin 100 tuhatta vuotta sitten, eli aika-asteikamme mukaan vain 31. joulukuuta viimeisen 20-15 minuutin aikana, ja ihmiskulttuurin historia muinaisesta Egyptistä nykypäivään kestää vain viimeiset 3 -5 minuuttia vuodesta!
Sointujen kehityksen kronologia saa meidät kiinnittämään huomiota evoluutioprosessin epätasaisuuteen, jossa voimakkaan muovaamisen jaksot korvattiin suhteellisen hitaiden ja kapeiden adaptiivisten muutosten aikoilla. A. N. Severtsov ehdotti tällaisen säännöllisen vuorottelun nimeämistä aromorfisten evoluutiomuutosten jaksojen muutokseksi, joka johtaa morfofysiologisiin muutoksiin, jotka nostavat organismin elintärkeän toiminnan korkeammalle energiatasolle (vitaalitoiminnan energian kasvu) - idioadaptaatiojaksoja tai saavutettujen etujen toteutuminen lisäämällä lukumäärää, laaja asutus, sopeutuminen paikallisiin olosuhteisiin ja hajoaminen alaryhmiin (perheet, suvut, lajit). G. Osborne kutsui viimeistä prosessia mukautuvaksi säteilyksi. Akordaattien muodostuminen ja evoluutio on klassinen esimerkki aromorfisesta polusta - luokkien syntymisestä - ja kunkin niistä seuraava idioadaptiivinen kukinta, joka puolestaan omisti hallitsevan aseman yksittäisten ajanjaksojen eläimistössä.
On syytä kiinnittää huomiota vielä yhteen sointujen kehityksen piirteeseen. Uusi aromorfoosien muodostama luokka, yleensä kehityksensä alkuvaiheessa, erotti ala-asemassa olevia oksia, jotka usein työntyivät takaisin epäsuotuisiin biotooppeihin. Pakko hallita uusia ympäristöjä ja sopeutua vieraisiin elinoloihin, ne kehittyivät hitaasti, mutta saattoivat saada yleistä merkitystä mukautuksia ja syntyessään ja vakiinnuttuaan pakottivat edeltäjänsä pois. Yleensä tätä edelsi muutos maan pinnassa (vuoristorakentamisen syklit), ilmasto ja kasvillisuus.
Niin, Agnatha ja Gnathostomata muodostui ilmeisesti lähiaikoina, mutta sitten leuattomat, jotka sopeutuivat elämään virtaavissa vesissä, pakottivat leuattomat ulos useimmista ekologisista markkinaraoista. Sama tapahtui kalojen evoluutiossa: panssaroidut kalat korvattiin rustoisilla kaloilla ja jälkimmäiset luisilla kaloilla. Kun maan selkärankaiset (sammakkoeläimet) ilmestyivät, niiden muinaisesta ryhmästään (ichthyostegs) erottui varhain haara, joka myöhemmin synnytti matelijoita, ja jälkimmäisten muodostumisen alkuvaiheessa ryhmä eläinhammaseläimiä erottui, mikä synnytti nisäkkäille. On korostettava, että uuden sointuluokan muodostuminen on aina liittynyt uuden "sopeutuvan vyöhykkeen", uuden elinympäristön kehittymiseen (G. Simpson). Joten hordaateista, jotka lähtivät merestä makeisiin vesiin, tuli selkärankaisia; evoluutio ja muutokset kalaluokissa liittyivät peräkkäiseen tunkeutumiseen suiston vesiin ja jokien alajuoksulta yläjuoksulle. Tämä on vielä ilmeisempi tetrapodien (maanpäällisen) selkärankaisten superluokassa. Evoluution olosuhteita, tekijöitä ja polkuja tarkastellaan yksityiskohtaisemmin kuvattaessa yksittäisiä selkärankaisten luokkia.
Selkärankaisten evoluution aikana ei vain rakenne (kehoorganisaatio) kehittynyt asteittain, vaan hermoston ja toiminnan kehityksen perusteella yksilöiden suhteet monimutkaistuvat ja populaatioorganisaation merkitys kasvoi. Monimutkaista ja tilavaa tietoa optisten, akustisten, kemiallisten ja muiden kanavien kautta välittävien viestintävälineiden monimutkaisuus varmistaa tehokkaan lisääntymisen, virtaviivaistaa eläinten alueellista jakautumista, parantaa avaruudessa suuntautumista ja lisää ympäristövaikutuksia. Liikkuvat ryhmittymät (perheet, karjat ja parvet) laajentavat luonnonvarojen käyttömahdollisuuksia ja lisäävät mahdollisuuksia olemassaolotaistelussa.
Viitteet: Naumov N.P., Kartashev N.N. Vertebrate Zoology. - Osa 2. - Matelijat, linnut, nisäkkäät: oppikirja biologille. asiantuntija. Univ. - M.: Korkeampi. koulu, 1979. - 272 s., ill.
Selkärankaisten elinjärjestelmät
| Parametrin nimi | Merkitys |
| Artikkelin aihe: | Selkärankaisten elinjärjestelmät |
| Otsikko (teemaattinen luokka) | Ekologia |
| Elinjärjestelmä | Järjestelmän toiminnot | Elimet, jotka ovat osa järjestelmää |
| kehon sisäosat | Kehon osat erottavat eläimen kehon ulkoisesta ympäristöstä ja suojaavat sitä haitallisten tekijöiden vaikutuksilta. Samalla ne eivät anna eläimen kehon antaa tarvitsemiaan aineita ulkoiseen ympäristöön. Monilla eläimillä kehon iholla on tärkeä rooli kaasunvaihtoprosesseissa. | Nahka |
| Tuki- ja liikuntaelimistön | Eläimen tuki- ja liikuntaelimistö varmistaa sen liikkumisen avaruudessa. Se on myös eläimen kehon runko ja auttaa sitä säilyttämään tietyn muodon. Toinen tuki- ja liikuntaelinten tehtävä on suojaava - se suojaa sisäelimiä mekaanisilta vaurioilta. | Luusto ja lihakset |
| ruoansulatus- | Ruoansulatusjärjestelmä tarjoaa elimistölle ravinteita. Se käsittelee orgaanisia aineita, jotka tulevat kehoon ulkoisesta ympäristöstä. | Hampaat, kieli, sylkirauhaset, ruokatorvi, maha, suolet, maksa, haima |
| Kuljetus | Kuljetusjärjestelmä varmistaa aineiden nopean siirtymisen kehon osasta toiseen. Sillä on myös tärkeä rooli monien prosessien säätelyssä, koska se on hormonien, ravintoaineiden ja aineenvaihduntatuotteiden kantaja. | Sydän, verisuonet, suonet, kapillaarit |
| Hengitys | Hengityselimet kuljettavat happea elimistöön ja poistavat siitä hiilidioksidia. | Keuhkot, kidukset, iho |
| erittäviä | Eritysjärjestelmä poistaa aineenvaihduntatuotteita eläimen kehosta. | Munuaiset, keuhkot, iho, virtsanjohdin, virtsarakko |
| endokriininen | Endokriiniset järjestelmät osallistuvat kehon yksittäisten elinten ja elinjärjestelmien toiminnan säätelyyn. Se tarjoaa elimistölle pitkäaikaisia reaktioita muuttuviin olosuhteisiin. Biologisesti aktiivisia aineita - hormoneja - käytetään välittämään signaaleja endokriinisessä järjestelmässä. | Hypotalamus, aivolisäke, haima, sukurauhaset, kilpirauhanen, käpylisäke |
| hermostunut | Hermosto tulkitsee aistielinten merkit ja koordinoi niiden kanssa kaikkien muiden elinten toimintaa. Se osallistuu myös kehon yksittäisten elinten ja elinjärjestelmien toiminnan säätelyyn. Hermosto reagoi nopeasti muuttuviin olosuhteisiin | Aivot, selkäydin, hermot |
| tuntoelimet | Eläinten aistielimet havaitsevat ympäristöstä tulevaa tietoa ja välittävät sen hermostoon. | Kosketus-, haju-, maku-, kuulo-, näkö-, tasapainoelimet |
| lisääntymiskykyinen | Lisääntymisjärjestelmä varmistaa eläinten lisääntymisen | sukupuolielimiä |
3. Monisoluisten organismien toimintojen säätely.
Eläinten organismin toimintojen säätely on parasta tarkastella ihmisen esimerkissä. On olemassa kaksi perussäätelymenetelmää: hermosto (käyttämällä hermoimpulsseja, jotka välittyvät hermosolujen kalvojen kautta) ja humoraalinen (käyttäen eri kehon nesteiden kuljettamia kemikaaleja).
Humoraalinen säätely- kehon fysiologisten toimintojen koordinointi eri kehon nesteiden (veri, imusolmukkeet, kudosnesteet) kuljettamien kemikaalien avulla - hormonit. Endokriinisen järjestelmän tekemä.
Endokriininen järjestelmä- joukko elimiä, elinten osia ja yksittäisiä soluja, jotka erittävät hormoneja vereen ja imusolmukkeisiin. Yhdessä hermoston kanssa se säätelee ja koordinoi tärkeitä ihmiskehon toimintoja: kasvua, lisääntymistä, aineenvaihduntaa, sopeutumisprosesseja.
Endokriinisessa järjestelmässä erotetaan keskus- ja reunaosat, jotka ovat vuorovaikutuksessa keskenään ja muodostavat yhden järjestelmän. Endokriinisen järjestelmän tehokkaan toiminnan perusta on palauteperiaatteen käyttö.
Ihmiskehon prosessien hermosäätely tapahtuu somaattisten ja autonomisten hermostojen avulla.
Somaattinen hermosto koostuu niistä keskus- ja ääreishermoston osista, jotka hermottavat luurankolihaksia ja aistielimiä. Se tarjoaa keholle tiedon havaitsemisen ulkoisesta ympäristöstä sekä toimista (luurankolihasten eri liikkeiden muodossa) vastauksena ulkoisiin tekijöihin.
Somaattisen hermoston tarjoamat liikkeet toteutetaan yksittäisten motoristen yksiköiden (lihaskuituryhmien, joista jokaista hermottaa yksi motorinen neuroni) koordinoiduilla toimilla.
Autonominen (kasvillinen) hermosto- hermoston osa, joka säätelee sisäelinten, rauhasten, verisuonten, sileiden ja joidenkin poikkijuovaisten lihasten toimintaa ja hallitsee myös aineenvaihduntaprosesseja.
Autonominen hermosto koostuu kahdesta osasta, joilla on päinvastainen vaikutus kehon elimiin ja kudoksiin - sympaattisesta ja parasympaattisesta jaosta. Autonomisen hermoston korkein ohjauskeskus on hypotalamus, joka ohjaa myös endokriinisen järjestelmän toimintaa.
Autonominen hermosto tarjoaa hermotuksen sisäelimille, verisuonijärjestelmälle, rauhasille ja sileille lihaksille. Sillä on myös troofinen vaikutus luurankolihaksiin. Aiheuttamatta näiden lihasten supistumista se parantaa niiden ravintoa ja siten stimuloi heidän työtä. Se säätelee sisäelinten ja verisuonten toimintaa, rauhasten eritystä ja sydämen toimintaa. Aineenvaihduntaprosesseja säätelee myös autonominen hermosto.
Autonomisen hermoston toiminta ei ole ihmisen tahdon ja tietoisuuden alaista. Ihminen ei edes tunne monien sisäelinten läsnäoloa, varsinkaan sellaisia, jotka eivät liiku, kuten esimerkiksi rauhaset, ei tunne kuinka ne erittävät, kuinka ruoka imeytyy suolistossa ja niin edelleen. Ihminen ei voi tietoisesti hallita näiden elinten toimintaa, koska hän hallitsee lihaksiaan. Tällaiset prosessit tapahtuvat ihmisen tietoisuuden ulkopuolella eivätkä ole hänen tahdon alaisia.
Autonomisessa hermostossa, kuten somaattisessa hermostossa, on keskus- ja ääreisosia. Keskiosaa edustavat vegetatiiviset neuronit, jotka muodostavat kertymän aivoihin ja selkäytimeen - vegetatiivisiin ytimiin. Perifeerinen osa muodostuu lukuisista autonomisista hermosolmukkeista ja hermosäikeistä.
Selkärankaisten elinjärjestelmät - käsite ja tyypit. Luokan "Selkärankaisten elinjärjestelmät" luokitus ja ominaisuudet 2017, 2018.
Selkärankaisten järjestö
Selkärankaisia yhdistää yhteinen morfofysiologinen organisaatio. Näiden eläinten kaikissa elinjärjestelmissä voidaan jäljittää peräkkäisten muutosten piirteitä homologisten elinten evoluutiomuutoksen yhteydessä. Alla on yleissuunnitelma yksittäisten elinjärjestelmien rakenteesta, toiminnasta ja rakenteesta.
Iho edustavat erittäin tärkeää toiminnallista järjestelmää. Iho on suorassa kosketuksessa ulkoisen ympäristön kanssa ja vaikuttaa suoraan siihen. Iho ja lihakset muodostavat eläimen kehon pinnasta, antavat sille muodon ja pitävät kaikki sisäelimet. Ihosuojukset suojaavat kehoa ulkoisilta mekaanisilta ja kemiallisilta vaurioilta, lämpöaltistumiselta, kuivumiselta ja mikrobien tunkeutumiselta. Iho osallistuu lämmönsäätelyyn, kaasunvaihtoon ja hajoamistuotteiden erittymiseen. Ihon johdannaiset voivat osallistua liikuntaelinten (sorkat) muodostumiseen, tarttua (kynnet), hyökkäämiseen ja puolustamiseen (sarvet, neulat jne.), lentoon (laskokset), uimiseen (verkot).
Iho sisältää kosketuselinten reseptoreita, siinä on monia rauhasia eri tarkoituksiin (lima-, rasva-, haju-, hiki- jne.).
Selkärankaisten iholle on ominaista kaksi kerrosta. Ulkokerros - orvaskesi - on ektodermaalista alkuperää. Se on aina monikerroksinen. Sen alempi kerros pysyy elossa ja aktiivisena koko elämän ajan ja tuottaa uusia solukerroksia. Epidermiksen ylemmät kerrokset koostuvat yleensä litteistä soluista, jotka maan selkärankaisilla keratinisoituvat, kuolevat ja jatkuvasti
räjähtää pois. Orvaskesta muodostuu kiimainen ihon johdannainen - kiimainen suomu, höyhenet, hiusraja, kynnet, sorkat, ontot sarvet. Epidermiksissä kehittyy erilaisia ihorauhasia.
Ihon sisäinen kuitukerros corium, jota kutsutaan muuten nimellä cutis, tai iho itse, kehittyy mesodermaalisesta rudimentista - somiitin iholehdestä. Corium-kerros on paksu, se muodostaa pääosan ihosta, jolla on suuri lujuus. Coriumissa kehittyy erilaisia luutumia kalan suomuina, sisäluuina, jotka muodostavat niin sanotun ihorungon (toisin kuin kondraalisessa luurangossa). Itse ihon kustannuksella peuran luusarvet kehittyvät myös. Kynsän alaosaan kertyy ihonalainen rasvakerros.
lihaksisto. Ihon alla sijaitseva lihaskerros on suurin osa lihaksistosta, ns kehon lihaksisto, tai somaattinen. Se tarjoaa eläimille mahdollisuuden liikkua ympäristössä ja koostuu poikkijuovaisesta lihaskudoksesta. Alemmilla selkärankaisilla, kuten ei-kalloisilla, lihaksisto on segmentoitunut. Korkeammissa selkärankaisissa kehon liikkeiden yleisen komplikaatioiden vuoksi raajojen kehittyessä segmentointi häiriintyy ja rungon lihakset ryhmitellään muodostaen sellaisia kehon osia kuin vartalo, pää ja liikeelimet.
Paitsi somaattiset lihakset Selkärankaisilla on suoliston lihakset ja jotkut muut sisäelimet (suonet, kanavat). Tätä lihasta kutsutaan viskeraaliksi. Se koostuu sileästä lihaskudoksesta ja tarjoaa erityisesti ruoan liikkumisen suolistossa, verisuonten seinämien supistumista.
Vartalon lihakset alkavat alkiona myotomin sisäkerroksesta (katso lansetin kehitys), ts. dorsaalinen mesoderma. Viskeraalinen lihaksisto on sivulevyn johdannainen, ts. vatsan mesoderma.
Sisäinen luuranko- Tämä on selkärankaisen eläimen kehon tukipohja. Lisäksi luuranko on mukana kehon liikkeessä, suojaa sisäelimiä.
Topografisesti selkärankaisten luuranko voidaan jakaa aksiaaliseen, viskeraaliseen, raajavyön ja vapaan raajan luurankoihin.
Aksiaalinen luuranko alkuperäisessä muodossaan esitetään sointu jota ympäröi paksu sidekudoskalvo. Jälkimmäinen ei kata vain sointua, vaan myös sen yläpuolella olevan hermoputken. Notochord kehittyy primaarisen suolen dorsaalisen puolen alkeesta, ts. on endodermaalista alkuperää.
Riisi. 9. Selkärangan muodostuminen (poikittaisleikkaukset):
1 - alemman kaaren alku; 2 - yläkaaren alku; 3 - kylkiluu, 4 - hemaalinen prosessi, 5 - nikaman lateraalinen prosessi, 6 - yläkaari
Useimmissa selkärankaisissa notochord siirtyy ja korvataan rustoisella tai luisella luurangolla. Rusto- ja luurungot kehittyvät edellä mainitun sidekudoskalvon (alkuperäinen mesodermi) kalvon johdannaisina. Tämä kuori on siis luuranko.
Aksiaalisessa luurangossa on selkäranka ja aivokallo. Selkänikamien kehityksen aikana asetetaan aluksi metameerisesti sijoitetut parilliset rustot notochordin pinnan viereen. Nämä ovat ylemmän ja alemman nikamakaaren alkeet (kuva 9.1). Yläkaarien ulkopäiden kasvu ja sulkeminen johtaa muodostumiseen selkärangan kanava, jossa hermoputki sijaitsee (kuva 9, II). Alemmat kaaret sulkeutuvat hännän alueella (kaloissa) ja rajoittavat hemal kanava, mistä selkä-aortta ja häntälaskimo kulkevat. Ylä- ja alakaaren sisäpäiden sulkemisen seurauksena nikamakappaleet muodostuvat, niiden sisällä ja välissä voi jossain määrin säilyä notokordi (kuva 9, III). Vartalon alueella kylkiluut on kiinnitetty alempien nikamakaarien prosesseihin.
aivokallo kallo on asetettu kahteen rustopariin, jotka sijaitsevat aivojen alkupään alla. Heidän takaparinsa - parachordalia- sijaitsee jänteen etupään sivuilla; etupari - trabeculae- hänen edessään. Ne muuttavat ensimmäisten nikamien alkioita ja osittain sisäelinten luurangon kaaria. Parachordalian ja trabeculien kasvu ja sulkeutuminen johtavat kallon päälaminan muodostumiseen, joka on aivojen alla. Samaan aikaan muodostuvien aistielinten (haju, näkö, kuulo) ympärille ilmestyy rustoisia kapseleita. Ne sijaitsevat hieman päätason yläpuolella

Riisi. 10. Hain rustokallon kehitys:
1 - kiduskaaret; 2 - hyoidikaari; 3 - leuan kaari; 4 - kuulokapseli; 5 - sointu; 6 - aivolisäke; 7 - suolisto; 8 - keskiaivot; 9 - selkäydin; 10 - sierain; 11 - silmä; 12 - parachordalia; 13 - primaarinen etuaivot; 14-orbitaaliset rustot; 15 - trabekulaat
kallon levyt ja peittävät aivot sivuilta. Myöhemmässä kehityksessä aistielinten kapselit yhdistetään ja sulautuvat kallon päälamelliin. Kallon rustotilassa aivokotelossa ei esiinny täyttä kattoa. Kallon katon kammien väliin jääneet reiät - fontanelles- Kiristää sidekudoskalvolla. Kallon kiinteä katto esiintyy vain päällekkäisten (iho)luiden (etu-, parietaali) muodostumisen yhteydessä.
Siten aivokallo syntyy aivojen ja aistielinten kehityksen yhteydessä niiden suojaavana muodostelmana.
Viskeraalinen luuranko fylogeneettisesti muodostunut kallosta riippumatta (kuva 10). Sen muniminen tapahtuu sidekudoksessa lähellä ruoansulatusputken etuosaa. Aluksi viskeraalinen luuranko on sarja lukuisia yksitoikkoisia kaaria, jotka sijaitsevat kidusrakojen välissä. Ne toimivat hengityslaitteiden tukena.
Myöhempi viskeraalisen luuston muutos liittyy selkärankaisten sellaisten elinten hankkimiseen, kuten ylä- ja alaleuat, välikorva, aivokallon pohja ja kurkunpää.
Käsitettä "viskeraalinen luuranko" tarkastellaan pääasiassa alempien selkärankaisten yhteydessä. Korkea-asteen koulutuksessa se korvataan käsitteillä "viskeraalinen kallo", "kasvojen kallo".
Raajavyön luuranko ja vapaat raajat. Selkärankaisilla erotetaan parittomat ja parilliset raajat. Parilliset raajat puolestaan voivat olla joko eviä tai maaperäisiä raajoja.
Parittomien raajojen luuranko - selkä-, hännän-, peräevät- koostuu useista rusto- tai luusäteistä, jotka eivät ole yhteydessä luuston muihin osiin.
Parillisten raajojen luuranko on jaettu raajavöiden luurankoon ja vapaan raajan luurankoon. Raajavyöt sijaitsevat aina eläimen kehon sisällä. Selkärankaisten vapaata raajan luurankoa on kahta tyyppiä: kalaevä ja

Riisi. 11. Kaavio selkärankaisten luurangoista:
I - kalan luuranko, II - maan selkärankaisen luuranko
viisisormeinen raaja maan selkärankaiset. Ensimmäisessä tapauksessa luurankoa edustavat useat rustojen tai luiden rivit, jotka liikkuvat vyön suhteen yhtenä vipuna. Viisisormen raajan luuranko koostuu useista vipuista, jotka voivat liikkua sekä yhdessä suhteessa raajan vyöhön että erikseen - toinen suhteessa toiseen (kuva 11). Raajan luuranko laskeutuu ihon sidekudoskerroksessa.
Ruoansulatuselimet. Ruoansulatusjärjestelmää edustaa putki, joka alkaa suusta ja päättyy peräaukkoon. Ruoansulatuskanava muodostuu gastrulan endodermaalisesta putkesta (katso lansetin kehitys). Tässä suhteessa ruoansulatuskanavan epiteeli on endodermaalinen. Vain suun ja peräaukon aukkojen alueella endodermaalinen epiteeli siirtyy huomaamattomasti ektodermaaliseen. Tämä johtuu kehon seinien (ja siten ektodermin) tunkeutumisesta yllä olevien reikien muodostumisen aikana.
Ruoansulatuskanava on jaettu seuraaviin pääosiin: 1) suuontelon tarjoilu syömiseen; 2) nielu- osasto, joka liittyy aina hengityselimiin: kaloilla kidusraot avautuvat nieluun, maaselkärankaisilla kurkunpään rako sijaitsee nielussa; nielua kutsutaan oikeutetusti ruoansulatusputken hengitysosastoksi; 3) ruokatorvi, 4) vatsa- suoliston laajeneminen, jossa joissakin tapauksissa on erittäin monimutkainen laite; 5) suolet, tyypillisesti jaettu etu- tai ohueen, keski- tai paksuun ja taka- tai peräsuoleen. Suolikanavan morfologinen komplikaatio selkärankaisten sarjassa seuraa polkua
sen venyminen ja eriyttäminen osastoihin. Kolmen tyyppisen ruoansulatusrauhasen kanavat avautuvat ruuansulatusputkeen: sylki, maksa, haima.
Sylkirauhaset- Maan selkärankaisten hankinta. Ne muuttavat suuontelon limakalvoja. Niiden salaisuus kostuttaa ruokaa ja edistää hiilihydraattien hajoamista.
Maksa ja haima kehittyvät alkiosuolen etuosan ulkonemalla. Maksa syntyy suolen vatsan seinämän sokeasta kasvusta (katso lansetin suolen maksakasvu). Maksakanavat valuvat ohutsuolen etuosaan. Haima kehittyy useasta, useammin kolmesta alkeesta, jotka ovat myös suolen kasvaimia. Tämä rauhanen, toisin kuin maksa, ei yleensä näytä kompaktilta rungolta, ja sen lohkot ovat hajallaan ohutsuolen etuosassa.
Molempien näiden rauhasten toiminnot ovat laajempia kuin vain ruoansulatus. Maksa on siis sen lisäksi, että se erittää sappia, emulgoi rasvoja ja aktivoi muiden ruoansulatusentsyymien toimintaa, ja se on tärkeä aineenvaihduntaelin. Täällä jotkut haitalliset hajoamistuotteet neutraloituvat, glykogeeni kerääntyy. Haimaentsyymit hajottavat proteiineja, rasvoja ja hiilihydraatteja. Samaan aikaan haima toimii sisäisen erityksen elimenä. Tämän toiminnon hajoaminen johtaa elimistön kyvyn käyttää sokeria menetykseen. Tuloksena on vakava sairaus - diabetes.
Hengitysjärjestelmä Selkärankaisia on kahta tyyppiä - kidukset ja keuhkot, ja merkittävällä osalla selkärankaisista iho on välttämätön hengityksen kannalta.
Kiduslaite on järjestelmä pareittain, yleensä symmetrisesti sijaitsevista rakoista, jotka viestivät nielusta ulkoisen ympäristön kanssa. Kidusten rakojen etu- ja takaseinämät on vuorattu limakalvolla, joka muodostaa lamellikasvuisia; kasvatukset jaetaan terälehtiä, jossa on nimi kidukset. Jokainen terälehtien yläpuolella oleva kiduslevy on nimetty puolikidukset. Kidusten välisissä väleissä (kidusten väliseinäissä) hajoavat sisäelinten kiduskaaret(Katso s. 27 sisäelinten luuranko). Siten kukin kidusten kaari on yhdistetty kahteen kidusten puolikkaaseen kahdesta erilaisesta kidusraosta.
Kiduksen halkiot on asetettu parillisten endodermaalisten ulkonemien järjestelmän muodossa, jotka kasvavat ulospäin nielusta. Samaan aikaan ulomman ihon ektodermaalisia invaginaatioita ilmaantuu. Primordiat kasvavat toisiaan kohti ja sitten yhdistyvät. Siksi kidusraot ovat sekoitettuja ento- ja ektodermaalista alkuperää. Filamentit Yleensä
kehittyvät halkeaman ektodermaalisesta alkeesta, ja vain leuattomilla eläimillä ne ovat endodermaalista alkuperää.
Maan selkärankaisten hengityselimet - keuhkoihin- kaaviossa ne ovat pussipari, jotka avautuvat nieluun kurkunpään halkeaman kautta. Alkiossa keuhkot syntyvät nielun vatsan seinämän ulkonemana kiduksen takaosassa, ts. ovat endodermaalista alkuperää. Alkion kehityksen alkuvaiheessa keuhkosymut muistuttavat sisäisiä (endodermaalisia) kidusten rakoja. Nämä olosuhteet sekä keuhkoille ja kiduksille yhteiset verenkierto- ja hermotusominaisuudet pakottavat meidät pitämään keuhkoja homologeina takimmaisen kiduspussin parin kanssa.
Nahka osallistuu hengitykseen tapauksissa, joissa siinä ei ole tiheitä kiivaisia tai luisia suomuja, esimerkiksi sammakkoeläimillä, paljasnahoisilla kaloilla.
Hengityselimet osallistuvat toiminnallisesti veren rikastamiseen hapella ja hiilidioksidin poistoon. Ammoniakkia vapautuu alempien vesieläinten hengityselinten kautta. Lämminverisillä eläimillä se osallistuu lämmönsäätelyprosesseihin. Hengitysjärjestelmän toimintaperiaate on CO 2:n ja O 2:n vaihto kaasun ja veren virtausten välillä, jotka on suunnattu vastavirtaan toisiaan kohti.

Riisi. 12. Kalojen (I) ja selkärankaisten (II) verenkiertokaavio:
1 - haaravaltimot; 2 - kaulavaltimo; 3 - dorsaalinen aortta, 4 - vatsa-aortta; 5 - keuhkovaltimo; 6 - Cuvier-tiehy, 7 - etupään (kaulalaskimo); 8 - posterior kardinaalilaskimo; 9 - posterior vena cava; 10 - maksalaskimo; 11 - keuhkolaskimo 12 - maksan porttilaskimo; 13 - suolenalaiseen laskimoon
Verenkiertoelimet. Verenkiertoelimet selkärankaisilla, kuten ei-kalloilla, on suljettu. Verenkiertojärjestelmä asetetaan sivulevyjen sisälevyistä (katso lansetin kehitys). Se koostuu toisiinsa yhdistetyistä verisuonista, jotka voidaan karkeassa kaaviossa vähentää kahdeksi rungoksi: selkä- missä veri virtaa päästä häntään ja vatsan- jota pitkin se liikkuu vastakkaiseen suuntaan. Toisin kuin ei-kallo (kuva 12), selkärankaisilla veren liike liittyy sydämen toimintaan.
Sydän on paksuseinäinen lihaksikas pussi, joka on jaettu useisiin osiin - kammioihin. Sydämen pääosat ovat eteis, joka vastaanottaa verta, ja kammio, joka ohjaa sitä kehon läpi. Sydänkammioiden lukumäärä on erilainen eri selkärankaisten luokissa.
Alkiossa sydän syntyy vatsa-aortan takaosan laajenemisena, joka tässä paikassa kiertyy kaarevaksi silmukaksi. Silmukan etuosa synnyttää kammio sydämet, selkä - atrium.
Sydämessä on poikkijuovaiset lihakset, jotka toimivat automaattitilassa, eikä sen supistuminen ole tahdonvoimaisten impulssien alainen. Sydämen mitat liittyvät sen työn intensiteettiin, ja sen mitat suhteessa kehon mittoihin kasvavat selkärankaisten sarjassa (taulukko 1).
pöytä 1
Sydänindeksi eri luokkien selkärankaisilla
Verisuonet on jaettu kahteen järjestelmään: valtimoiden jossa veri virtaa sydämestä ja laskimo jonka kautta veri palaa sydämeen. Selkärankaisten komplikaatioprosessissa tapahtuu siirtymä eläimistä, joilla on yksi verenkiertokierto, kahden verenkiertopiirin omistajiin.
Luonteeltaan veri tarkoittaa sidekudosta, joka tunkeutuu verenkiertoon solujen välisestä tilasta.
Selkärankaisten veri koostuu värittömästä nesteestä - plasmasta, jossa muodostuneet veren elementit sijaitsevat: punasolut tai punasolut, jotka sisältävät väriainetta - hemoglobiinia, ja valkosolut - leukosyytit. Punasolut liittyvät veren hapettumiseen, koska ne kuljettavat happea. Leukosyytit osallistuvat elimistöön joutuneiden mikro-organismien tuhoamiseen. Lisäksi veressä on verihiutaleita, joilla on tärkeä rooli veren hyytymisessä, samoin kuin muissa soluissa. Veren massa selkärankaisten evoluutiosarjassa kasvaa (taulukko 2).
taulukko 2
Veren suhteellinen massa kehon massaan eri luokkien selkärankaisilla
(Prosserin ja Brownin, 1967; Prosserin, 1978 jälkeen)
Verenkiertojärjestelmä on monikäyttöinen. Se osallistuu elinten, kudosten, solujen hapen, orgaanisten ja kivennäisaineiden, nesteiden vastaanottamiseen ja hajoamistuotteiden, toksiinien, hiilidioksidin poistamiseen, hormonien siirtoon endokriinisissä rauhasissa jne.
Verenkiertojärjestelmän ohella selkärankaisilla on siihen liittyvä toinen verisuonijärjestelmä - lymfaattinen. Se koostuu imusuonet ja imusolmukkeet. Imfaattinen järjestelmä ei ole suljettu. Vain sen suurilla suonilla on itsenäiset seinät, kun taas niiden haarat avautuvat eri elinten solujen välisiin tiloihin. Imusuonet sisältävät väritöntä nestettä - imusolmukkeet, jossa imusolmukkeisiin muodostuneet lymfosyytit uivat. Immun liike määräytyy seinien, suurten suonten joidenkin osien (ns. lymfaattisten sydämien) supistumisen ja ajoittain muuttuvan paineen perusteella eri elinten suonissa.
Lymfaattinen järjestelmä toimii välittäjänä aineiden vaihdossa veren ja kudosten välillä.
Hermosto. Hermoston toimintoja ovat ulkoisten ärsykkeiden havaitseminen ja esiin tulevien viritteiden välittäminen soluihin, elimiin, kudoksiin sekä yksittäisten elinjärjestelmien ja koko kehon toiminnan yhdistäminen ja koordinointi yhdeksi toimivaksi eläväksi. järjestelmä. Alkiossa selkärankaisten hermosto syntyy, aivan kuten ei-kalloistenkin, onton putken muodossa, joka on asetettu ektodermiin alkion selkäpuolelle (kuva 13). Myöhemmin sen erilaistuminen tapahtuu, mikä johtaa: a) keskushermosto, jota edustavat aivot ja selkäydin, b) ääreishermosto, joka koostuu aivoista ja selkäytimestä ulottuvista hermoista, ja c) sympaattinen hermosto, joka koostuu selkärangan lähellä sijaitsevista hermosolmukkeista, jotka on yhdistetty pitkittäisillä säikeillä.
Selkärankaisten aivoja edustaa viisi osaa: etu-, väli-, keski-, pikkuaivot ja pitkulaiset aivot. Se asettuu alkioon hermoputken etuosan turvotuksen muodossa, joka pian jakautuu kolmeen primääriseen aivorakkulaan (kuva 14). Tulevaisuudessa ensimmäinen aivorakko synnyttää etuaivojen etuosan; sen takaosa muuttuu välilihakseksi. Väliaivot muodostuvat toisesta aivorakosta. Ulkoneman avulla

Riisi. 13. Selkärankaisten keskushermoston peräkkäiset kehitysvaiheet (kaaviolliset poikkileikkaukset):
ektodermin I-II erilaistuminen; III hermolevyn invaginaatio, IV-V - hermoputken eristäminen, 1 orvaskesi; 2 gangliolevyä

Riisi. 14. Aivojen osien kehitys (kaavio):
I - kolmen kuplan vaihe (silmäkuplien kanssa), II - viiden osaston vaihe (silmäkupit), 1 - etuaivot, 2 - välilihas; 3 - keskiaivot; 4 - pikkuaivot; 5 - medulla oblongata; 6 - silmäkuppi; 7 - silmäkupla
kolmannen aivorakon katto muodostaa pikkuaivot, joiden alla sijaitsee pitkulainen ydin. Etuaivot jaetaan edelleen vasempaan ja oikeaan puoliskoon.
Samanaikaisesti hermoputken pääosan kasvun ja erilaistumisen kanssa tapahtuu vastaava neurocoelin muutos. Sen kaksi pidennystä etuaivojen pallonpuoliskolla tunnetaan nimellä sivukammiot aivot. Aivojen väliosassa olevaa neurocoelin laajennettua osaa kutsutaan nimellä kolmas kammio, keskiaivoontelo - as Sylvian akvedukti, ontelo pitkittäisytimen - as neljäs kammio, tai rombinen kuoppa(Kuva 14). 10 tai 12 paria poistuu aivoista aivohermot.
Etuaivoissa on edessä kaksi symmetrisesti sijoitettua ulkonemaa, joista ensimmäinen päähermopari lähtee - haju-(I). Välipään pohjasta lähde visuaalinen hermot (toinen päähermopari, II).
Välikefalonin katolle kehittyy kaksi jaloilla istuvaa ulkonemaa: etu - parietaalinen elin ja takaisin - epifyysi.
Välikefalonin pohjasta kasvaa pariton ulkonema - suppilo, joka on rakenteeltaan ja toiminnaltaan monimutkaisen muodostelman vieressä - aivolisäke. Etummainen aivolisäke kehittyy suuontelon epiteelistä, takaosa - medullasta. Se sijaitsee siellä hypotalamus.
Väliaivojen katto muodostaa parillisia turvotuksia - visuaalisia lohkoja (tubercles). Kolmas aivohermopari lähtee keskiaivoista ( okulomotorinen, III). Neljäs päähermopari ( lohko, IV) lähtee keskiosan ja pitkittäisytimen väliseltä rajalta, kaikki muut päähermot (V - X - XII) lähtevät ytimestä.
Selkäydin ei jyrkästi rajattu ytimestä. Selkäytimen keskellä (elimen pääakselia pitkin) on säilynyt neurocoel, joka tunnetaan selkärankaisilla nimellä selkärangan kanava.
Selkäydinhermot lähtevät selkäytimestä metameerisesti (segmenttien lukumäärän mukaan). Ne alkavat kahdella juurilla: selkä - sensorinen ja vatsa - moottori. Nämä juuret sulautuvat pian selkäytimestä poistumisen jälkeen muodostaen selkäydinhermot, jotka sitten jakautuvat jälleen selkä- ja vatsahaaroihin.
tuntoelimet. Tämä elinryhmä syntyy alkion eri osien johdannaisina ja sen eri kehitysvaiheissa. Nämä ovat haju-, näkö-, kuuloelimet, vestibulaarilaitteisto, sivulinjan elimet, makuelimet, kosketuselimet, tietyt elimet, jotka havaitsevat maan magneettikentän, sähkökentät, lämpösäteily jne.
Hajuelimet. Uskotaan, että hajuaisti on yksi aivojen vanhimmista toiminnoista. Hajuelimet asettuvat ektodermin paksuuntumiseen samanaikaisesti hermolevyn kanssa. Samanaikaisesti muodostuu hajukapseleiden luuranko, jotka ovat osa aivojen kalloa. Aluksi hajukapselit kommunikoivat vain ulkoisen ympäristön kanssa ja niillä on ulkoiset sieraimet. Myöhemmin, maan olemassaolon vuoksi, sieraimet tulevat läpi.
näköelimet kuuluvat myös muinaisiin aistielimiin. Valoherkkä vastaanotto tapahtuu sointujen evoluution hyvin varhaisessa vaiheessa (muista lansetti) ja muodostuu alkion varhaisessa vaiheessa. Selkärankaisten näköelimet jaetaan parillisiin ja parittomiin. Molemmat ovat aivokalvon kasvua. Parilliset silmät asetetaan välikalvon lateraalisten osien kasvuna, parittomat - peräkkäin välilihan katossa (käpyrauhanen ja parietaalinen elin). Parillisten silmien asettamiseen liittyy visuaalisten kapseleiden muodostuminen niiden ympärille, jotka ovat osa aivokalloa (kuvat 15, 16).
kuuloelimet niillä on monimutkainen alkuperä selkärankaisissa. Evoluution varhaisin on sisäkorva, joka

Riisi. 15. Silmän kehityksen peräkkäiset vaiheet:
1 - aivokalvon; 2 - orvaskesi, joka muodostaa linssin; 3 - silmäkupin muodostumisen alku; 4 - silmäkupin jalka; 5 - linssi; 6 - sarveiskalvo; 7 - verkkokalvo; 8 - pigmenttikuori; 9 - mesodermisolut, jotka muodostavat suonikalvon ja kovakalvon

Riisi. 16. Ihmissilmän sagittaalinen osa:
1 - sarveiskalvo; 2 - iiris; 3 - etukammio; 4 - yksi silmää liikuttavista lihaksista; 5 - lasimainen runko; 6 - näköhermo; 7 - kovakalvo; 8 - suonikalvo; 9 - verkkokalvo; 10 - sädelihas; 11 - sädekehä
asetetaan alkion ektodermiin, syvenee reiän muodossa ja muotoutuu kuulovesikkeli makaa kuulokapselissa. Kuulovesikkeli on jaettu supistumisen avulla kahteen osaan. Yläosa muuttuu vestibulaarilaitteeksi. Se on tasapainoelin. Sen avulla voit tuntea kehon sijainnin Maan kolmiulotteisessa avaruudessa. Tämä ruumis on 3 puoliympyrän muotoinen kanava sisäkorvassa (kuvat 17, 18). Kuulorakkulan alaosa on itse sisäkorva - kuulopussi.
Keski- ja ulkokorva muodostuvat selkärankaisten ilmaantumisen myöhäisissä vaiheissa rantautumisen yhteydessä.
Sivulinjan elimet vain primaarisille vedessä eläville selkärankaisille ominaisia, myös munitaan ektodermiin. Ne ovat uria, jotka venyvät

Riisi. 17. Sisäkorvan peräkkäiset kehitysvaiheet:
I - kuuloplakoodi, II - fossa, III ja IV - rakkula osassa; V ja VI - puoliympyrän muotoisten kanavien muodostuminen; 1 - etanan alkuosa; 2 - puoliympyrän muotoinen kanava; 3 - endolymfaattinen kanava; 4 - pyöreä laukku; 5 - soikea pussi
pitkin pään sivuja ja pitkin vartaloa. Luiset suomut voivat peittää nämä urat tai olla peittämättä niitä. Sivulinjaelimet havaitsevat valon liikkeet ja veden värähtelyt värähtelylähteiden lähellä: virtausten nopeuden ja suunnan, oman kehon liikkeitä ja esineiden läsnäoloa eläimen tiellä vedessä. Ne ovat seisminen anturijärjestelmä.
makuelimet ne asettuvat endodermiin ja havaitsevat alueella kulutetun ruoan maun: makea, karvas, hapan, suolainen. Ne sijaitsevat makuhermillä suuontelossa.
tuntoelimet. Niissä ei ole aistisoluja, vaan ne ovat ihossa haarautuneita hermopäätteitä, jotka havaitsevat ympäristön esineitä koskettamalla.

Riisi. 18. Kalvoisen kuulolabyrintin kaavio:
1, 2, 3 - etu-, ulko- ja takapuoliympyrän muotoiset kanavat, 4 - endolymfaattinen tiehye, 5 - kuulotäplät, 6 - pyöreä pussi, 7 soikea pussi, 8 - otoliitit

Riisi. 19. Bowmanin kapseli, jossa on verisuonten sotku:
1 - afferentti verisuoni, 2 - efferentti verisuoni, 3 - Bowmanin kapseli, 4 - verisuonten vyyhti, 5 - primaarinen virtsa, 6 - munuaistiehy, 7 - munuaistiehyen (nefrostomi) suppilo, joka avautuu kehon onteloon

Riisi. 20. Selkärankaisten virtsatiejärjestelmän kaavio:
I (uros) ja II (naaras) - hait ja sammakkoeläimet, III (uros) ja IV (naaras) - matelijat ja linnut, V (uros) ja VI (naaras) - nisäkkäät, 1 - pronephros (pronephros); 2 - kivekset ; 3 - vas deferens, 4 - mesonephros (primaarinen munuainen), 5 - takasuoli, 6 - rakko, 7 - kloaka, 8 - suppilo, 9 - munasarja, 10 - Muller-kanava, 11 - lisäkives (jäljellä meson etuosaa) - nefros), 12 - vas deferens, 13 - metanephros (sekundaarinen munuainen), 14 sekundaarinen virtsanjohdin, 15 mesonephros rudiment, 16 munanjohdinta, 17 munasolua, 18 munanjohtimen seinämän rauhasten erittämää proteiinia, 19 - kohtu, 20 - alkio kohtu 21 - emätin, 22 - sukupuoliontelo, 23 - eturauhaset; 24 - siemenrakkula, 25 - vas deferens, 26 - epididymis, 27 - perineum; 28 - Muller-kanavan alkuosa, 29 - mesonephros-kanava (primaarinen munuainen); 30 - parituselin (penis), 31 - peräaukko
erityselimiä. Kaikilla selkärankaisilla on erityselimiä munuaiset suunniteltu poistamaan ylimääräistä vettä, mineraalisuoloja ja typen aineenvaihdunnan hajoamistuotteita urean tai virtsahapon muodossa elimistöstä. Ne ovat mesodermaalista alkuperää, ja ne sijaitsevat somiittien ulkoseinällä. Munuaisten rakenne ja toimintamekanismi eri selkärankaisten ryhmissä eivät kuitenkaan ole samat. Selkärankaisten evoluutioprosessissa kolmenlaisia munuaisia muuttuu: pää, tai pronephros (pronephros), runko, tai primaarinen, munuainen (mesonephros) ja lantio, tai toissijainen, munuainen (metanephros). Erilaisilla munuaisilla on erilaiset erittymisprosessin periaatteet: erittyminen kehon ontelosta, sekaeritys (ruumiinontelosta ja verestä) ja lopuksi vain verestä. Samaan aikaan tapahtuu muutoksia veden takaisinimeytymismekanismissa. Veden ja siihen liuenneiden proteiiniaineenvaihdunnan tuotteiden vapautuminen ruumiinontelosta on mahdollista monien ruumiinonteloon avautuvien suppilomaisten nefrostoomien ansiosta. Erittyminen verestä tapahtuu munuaisten Malpighian kappaleiden kautta. Munuaisten eritystiehyitä kutsutaan susi kanavat, ne vaihdetaan virtsanjohtimet. Useimmilla selkärankaisilla on virtsarakon. Ensisijaisessa vesieliössä ammoniakkia voidaan laskea kidusten läpi.
Sukupuolielimet. Selkärankaisten sukupuolirauhaset - naaraiden munasarjat ja urosten kivekset - ovat yleensä parillisia. Ne kehittyvät mesodermiosasta tämän rudimentin jakautumiskohdassa somiitiksi ja sivulevyksi.
Aluksi (leuattomissa) sukupuolirauhasissa ei ollut erityskanavia ja lisääntymistuotteet putosivat sukurauhasten seinämien repeämien kautta ruumiinonteloon, josta ne erittyivät ulkoiseen ympäristöön erityisten huokosten kautta. Myöhemmin syntyi sukuelimet, joka miehillä liittyy erityselimiin (Suden kanava). Ja naisilla se toimii munanjohtimina Muellerin kanava, joka ylläpitää kokonaisuuden yhteyttä ulkoiseen ympäristöön.
Selkärankaiset- lat. Selkärankaiset, näiden eläinmaailman edustajien ominaisuus on todellisen luun tai rustorungon läsnäolo, jonka perusta on selkäranka. Tämä ryhmä on hyvin monipuolinen ja sisältää seuraavat eläintyypit: kalat, sammakkoeläimet, matelijat, linnut ja nisäkkäät.
Selkärankaisten alkuperä ja eläintiede
Korkeammissa sointujen - selkärankaisissa tai kalloissa - notochord korvataan vähitellen nikamilla. Niiden luuranko on selkäranka. Selkärankaisilla on kallo. Heidän verenkiertoelimistöön kuuluu sydän. Toisin kuin alemmat chordatiryhmät, selkärankaiset etsivät ja sieppaavat aktiivisesti ruokaa. Tällainen aktiivinen ruokinta antoi selkärankaisille merkittäviä etuja olemassaolon taistelussa.
Selkärankaiset kehittyivät lansetin kaltaisista primitiivisistä sointuista, joita kutsutaan ensisijaiseksi ei-kalloiseksi.
Kalan alkuperä
Kalaluokan edustajat syntyivät noin 400 miljoonaa vuotta sitten. Nämä ovat heikoimmin organisoituneita selkärankaisia tutkimiemme eläinten joukossa. Samaan aikaan kalat ovat pitkän evoluution aikana sopeutuneet täydellisesti elämään vesiympäristössä. Aktiiviset liikeelimet, lihakset, sisäinen luuranko, kidukset, aivot ja aistielimet ovat saavuttaneet erityisen kehityksen.
Olennainen hetki kalojen kehityksessä oli parittomien evien muodostuminen. Tällä oli erittäin tärkeä rooli korkeampien selkärankaisten myöhemmässä evoluutiossa.
Eläinten kehityksen haara, joka johti maan selkärankaisten ilmestymiseen, erosi myös muinaisista kaloista.
Sammakkoeläinten alkuperä
Kauan ennen maan selkärankaisten ilmestymistä maapallolla asuivat muinaiset kalat - keilaeväkalat. Yksi näiden keilaeväkalojen haaroista, joka sopeutui maanpäälliseen olemassaoloon, synnytti ensisijaiset sammakkoeläimet. Siirtyessään elämään maalla, tällaiset crossopterygiat menettivät kiduksensa ja alkoivat hengittää keuhkoilla, evien sijaan he muodostivat viisisormeiset raajat, ja heidän verenkiertojärjestelmänsä muuttui paljon monimutkaisemmiksi.
Kuten eläinfossiilien tutkimus osoittaa, ensimmäiset sammakkoeläimet, jotka polveutuivat lohkoeväkaloista, ilmestyivät maapallolle yhden maapallon historian muinaisen aikakauden - devonikauden - kauden lopussa. Seuraavalla hiilikaudella sammakkoeläimet erottuivat jo lajien rikkaudesta ja monimuotoisuudesta ja saavuttivat laajan levinneisyyden.
Hiilikauden lämmin ja kostea ilmasto, jolle oli tunnusomaista poikkeuksellisen runsas maakasvillisuuden kehitys (puumaiset saniaiset, korteet ja sammalit), suosi sammakkoeläinten kehitystä. Maapallolla ei kuitenkaan tuolloin ollut korkeampia selkärankaisia (lintuja, nisäkkäitä).
Muinaiset fossiiliset sammakkoeläimet kuuluivat sukupuuttoon kuolleiden stegokefaalien ryhmään, joka tarkoittaa käännöksessä "kuoripäistä". Nämä eläimet saivat tämän nimen, koska niiden kallo oli ulkopuolelta peitetty kiinteällä kuorella. Kuori peitti osittain heidän ruumiinsa.
Monet kalojen merkit, jotka stegocephalians ovat säilyttäneet organisaatiossaan, toimivat suorana vahvistuksena niiden alkuperästä.
Yhtä vakuuttava todiste sammakkoeläinten alkuperästä kaloista ovat myös yhtäläisyydet nykyaikaisten sammakkoeläinten toukkien kalojen - nuijapäiden - kanssa: kalamainen vartalon muoto, hännän evä, kidusten hengitys jne.
Matelijoiden alkuperä
Yhdessä nykyaikaisten sammakkoeläinten tai sammakkoeläinten kanssa, stegokefaalit loivat perustan matelijoille. Matelijoiden ilmaantuminen liittyy läheisesti ilmaston muutoksiin, jotka tapahtuivat muinaisen aikakauden lopussa. Maapallon ilmasto kuivuu tällä hetkellä, vesistöt kuivuvat, kosteikot muuttuvat laajoiksi maa-alueiksi.
Tämä aiheutti asteittaisen muutoksen muinaisissa sammakkoeläimissä ja primaaristen matelijoiden syntymisen niistä.
Matelijoiden luokka on ensimmäinen todellisten maan selkärankaisten luokka.
Nopeasti kehittyneet matelijat ottivat pian hallitsevan aseman ja työnsivät sammakkoeläimet taustalle.
Matelijoiden erityisen nopea kehitys tapahtui seuraavalla - keskiaikakaudella. Juuri tähän aikaan, kuten tiedämme jo aiemmasta materiaalista, matelijat olivat valtavan kokoisia ja niitä edusti laaja valikoima lajeja. Ne olivat yleisiä maalla, vedessä ja ilmassa.
Fossiiliset matelijat ovat poikkeuksellisen kiinnostavia. Myöhemmin muinaisista matelijoista syntyivät ne eläinmaailman haarat, joista nykyisten matelijoiden lisäksi ovat peräisin myös linnut ja nisäkkäät.
Lintujen alkuperä
Nykyaikaisten lintujen muinainen esi-isä Archeopteryx (käännettynä "ensimmäiseksi siipiksi"), jonka rakenteeseen olemme jo perehtyneet, syntyi joidenkin matelijoiden sopeutumisesta lentoon.
Archeopteryxin löytö todistaa vakuuttavasti, että linnut polveutuvat matelijoista. Lintujen ja matelijoiden välistä suhdetta todistavat myös monet yhteiset piirteet niiden rakenteessa ja kehityksessä.
Archaeopteryxin rakenteen tutkimus osoittaa, että ensimmäiset linnut olivat vielä huonoja lentäjiä. Vasta myöhemmin, evoluutioprosessissa, koko organisaation asteittaisen parantamisen vuoksi, linnut sopeutuivat täydellisesti lentoon.
Nisäkkäiden alkuperä
Nisäkkäät ovat korkein selkärankaisten luokka, jotka ovat myös sukua matelijoille, nimittäin eläinhammasliskoille.
Vanhimmat nisäkkäiden fossiilijäännökset ovat olleet tiedossa keskiajan alusta lähtien.
Nisäkkäät erottuvat monista piirteistä, jotka ovat antaneet heille mahdollisuuden voittaa elämän taistelu. Tällaisia nisäkkäiden ominaisuuksia ovat hiusraja, joka suojaa niitä hyvin kylmältä, maitorauhaset sekä poikasten synnyttäminen kohdussa.
Nisäkkäille ominaista lämminverisyys antoi niille, kuten linnuille, suuria etuja olemassaolotaistelussa. Suuri merkitys nisäkkäiden progressiivisessa evoluutiossa on myös aivojen merkittävällä kehityksellä.
Keskiaika, jota joskus kutsutaan "Matelijoiden aikakaudeksi", korvattiin uudella aikakaudella, jota hallitsevat linnut ja nisäkkäät.
Niveljalkaisten evoluutio kambriassa
Devonista tunnetaan useiden maanpäällisten niveljalkaisten ryhmien edustajia: paleotsoinen panssaroitujen hämähäkkien ryhmä (Soluta), punkit (Acaromorpha) ja alemmat primaariset siivettömät hyönteisryhmät (Insecta). Epäilemättä maan selkärangattomien monimuotoisuus oli devonikaudella paljon suurempi kuin meille tulleet vähäiset fossiilijäännökset antavat ymmärtää.
Varhaisen hiilikauden toisella puoliskolla ilmestyi korkeampia hyönteisiä, joilla oli siivet, pterygootit. On todennäköistä, että siivekkäiden hyönteisten esi-isät siirtyivät kiipeilyelämään puiden rungoissa ja latvuissa. Liukumiseen pudotusten ja hyppyjen aikana nämä eläimet käyttivät kehon sivuttain litistettyjä ulkonemia väsyttävillä osilla. Näistä liikkumattomista lisäkkeistä mukautuvan evoluution prosessissa luonnonvalinta on muodostanut hämmästyttävän täydellisen lentokoneen. Aluksi siivet olivat rintakehän kaikissa kolmessa osassa, vaikka etupari säilytti edelleen alkeellisen rakenteen kiinteiden tasojen muodossa, kuten esimerkiksi Paleodictyoptera (Palaeodictyoptera) (kuva 48). Useimmat tunnetuista hyönteisryhmistä ovat menettäneet tämän etusiipiparin. Hiilen lopussa oli jo olemassa erilaisia lentävien hyönteisryhmiä, joista osa säilyi nykypäivään (maarfot, sudenkorennot, torakat, orthoptera, homoptera, skorpionit). Paleotsoisella kaudella hyönteiset olivat ainoat lentävät eläimet. Jotkut sudenkorennot saavuttivat sitten suuria kokoja: Ylähiilen - Ala-Permin Meganeuran rungon pituus oli 30 cm, siipien kärkiväli oli jopa 65 cm.
Maan selkärankaisten (tetrapodien) esi-isät olivat keilaeväkaloja, joilla oli kevyet ja voimakkaasti lihaksikkaat parilliset raajat - sarkopterygium, mikä luultavasti antoi näiden kalojen paitsi liikkua altaiden pohjaa pitkin tiheiden metsiköiden keskellä, myös päästä ulos matalaan. vettä, ryömi pieniä kanavia pitkin säiliöstä toiseen. Näiden raajojen sisäinen luuranko muodostui yhdestä suuresta luusta, joka oli nivelletty raajojen (olkapää- tai lantio-) vyön kanssa, jota seurasi kahden rinnakkaisen luun toinen osa, ja lukuisia pieniä luita sijaitsi distaalisesti (kuva 45, c). . Tässä järjestelmässä maaselkärankaisten raajojen luurangon prototyyppi on helposti nähtävissä.
Todennäköisesti jotkin lohkoeväkalalajet siirtyivät kehittämään uutta ravintopohjaa - runsasta selkärangattomien eläimistöä, joka asui matalissa vesissä, rannikon kasveissa ja vesistöjen rannoilla. Tämä mukautuvan evoluution suunta johti edelleen maaperän kehittymiseen nopeasti kehittyvine kasvistoineen ja eläimistöineen, joihin ei vielä kuulunut suuria petoeläimiä. Tämä ekologinen markkinarako oli maan selkärankaisten käytössä.
On huomattava, että sopeutumiset maaelämään kehittyivät itsenäisesti ja rinnakkain useissa keilaeväkalojen kehityslinjoissa. Tässä yhteydessä E. Jarvik esitti hypoteesin kahdesta eri lohkoeväkalaryhmästä (Osteolepiformes ja Porolepiformes) kuuluvien maaselkärankaisten difyleettisestä alkuperästä. Kuitenkin useat tiedemiehet (A. Romer, I. I. Shmalgauzen, K. Thomson, E. I. Vorobyeva) kritisoivat Yarvikin argumentteja. Nykyään useimmat tutkijat pitävät tetrapodien monofyleettistä alkuperää osteolepiformisista crossopteraneista todennäköisempänä, vaikka parafilian mahdollisuus on sallittu, ts. sammakkoeläinten organisoitumisen tason saavuttaminen useiden rinnakkain kehittyneiden osteolepiformisten kalojen läheisesti liittyvien fyleettisten linjojen avulla.
Sammakkoeläinten (luokka Amphibia) vanhimmat edustajat ichthyostegi (Ichthyostegalia, kuva 49) tunnetaan Grönlannin ylä-devonin esiintymistä. Ichthyostegit olivat melko suuria (yli 80 cm pitkiä) eläimiä, joilla oli jo hyvin kehittyneet maatyyppiset viisisormeiset raajat, joiden avulla ne pystyivät epäilemättä ryömiä maan yli. Ichthyostegit viettivät kuitenkin todennäköisesti merkittävän osan elämästään vesistöissä ja olivat Jarvikin osuvalla ilmaisulla "nelijalkaisia kaloja". Tämän todistaa niissä olevan todellisen hännän evä, joka muistuttaa lohkoeväkalojen häntää, sekä seismosensoristen reseptorien säilyminen aikuisen tilassa ("sivuviiva" - aistielimet, jotka toimivat vain vesiympäristössä , havaitsevat infraäänivärähtelyjä ja sähkömagneettisia kenttiä). Ichthyostegin kallossa oli säilynyt kidusten kannen luiden alkeet (mahdollisesti myös kiduksia oli kiinnitetty rustoisiin kiduskaareihin). Vartalo oli peitetty pienillä luisilla suomuilla.
Todennäköisesti vanhimmat sammakkoeläimet, kuten ichthyostegas, hävisivät vesiympäristössä kilpailun lukuisille ja monimuotoisille keilaeväkaloille, koska maatyyppiset parilliset raajat eivät voineet olla tehokkaita syvyysperäsintä uidessa. Viisisormeisten raajojen edut ilmenivät vain vesistöjen pienissä osissa ja maalla. Todennäköisimmin silmukkaeväkalojen ja niiden jälkeläisten - sammakkoeläinten - välillä oli devonilla kilpailusuhde, mikä vaikutti näiden ryhmien hajoamiseen erilaisiin sopeutumisvyöhykkeisiin: silmukkaeväkalat hallitsivat vesistöissä ja sammakkoeläimet miehittivät keskialueen. veden ja maan välissä olevat elinympäristöt (pieni uinti vaikeaa; kosteikot; liikakosteusalueet maassa). Ohiten huomautamme, että tämäntyyppinen elinympäristö, väli- ja raja-alue veden ja maan välillä, on säilynyt tähän päivään asti sammakkoeläimille edullisimpana sopeutumisvyöhykkeenä, jossa sammakkoeläimet ovat säilyttäneet etunsa sekä kaloihin että edistyneempiin maanpäällisiin nelijalkaisiin ryhmiin nähden. Myöhäisdevonin aikana vanhimpien sammakkoeläinten erilainen kehitys oli jo alkanut. Ichthyostegien lisäksi Devonikauden lopussa oli edustajia useista muista sammakkoeläinryhmistä, joiden fossiilisia jäänteitä löydettiin eri alueilla. On uteliasta, että sormien lukumäärä näissä eläimissä oli usein 7-8.
Hiilellä alkaa muinaisten sammakkoeläinten kukinta, jota edustavat ylemmällä paleotsoisella kaudella monenlaiset muodot (kuva 50), jotka on ehdollisesti yhdistetty stegokefaalien, ts. peitetyt päät. Tämä termi, jolla nyt ei ole taksonomista merkitystä, korostaa näiden eläinten ominaispiirrettä, jonka päätä suojasi ruisista peritty kiinteä luukuori (ns. tikattu kallo), jonka lävistivät vain sieraimien aukot. silmäkuopat ja "parietaalinen silmä".
Tunnetuimpia stegokefalian ryhmistä ovat labyrintodontit (Labyrinthodontia), joihin kuuluvat ichthyostegit. Heidän nimeensä liittyy omituinen hampaiden rakenteen piirre, joka on peritty myös keilaeväkaloista: kiille ja dentiini muodostavat lukuisia poimuja, jolloin poikittaisleikkaukseen tulee labyrinttiä muistuttava kuvio.
Labyrintodontit ylemmällä paleotsoisella kaudella olivat yksi yleisimmistä ja runsaimmista selkärankaisten ryhmistä. Mukana oli sekä pieniä että suuria, yli 1,5 metrin pituisia lomakkeita. Hiiltä hallitsivat heikot raajat ja pitkävartaloiset lajit, jotka todennäköisesti asuivat lukuisissa suoissa. Permikaudella ilmestyi suuria krokotiilin muotoisia stegokefaalia, joilla oli suuri litteä pää (samanlainen kuin Eryops, ks. kuva 50, b), sekä pieniä lajeja, joilla oli paremmin kehittyneet raajat ja lyhennetty runko ja häntä (Cacops, kuva 50, c). Nämä viimeksi mainitut asuivat todennäköisimmin pääasiassa maalla ja ruokkivat erilaisia selkärangattomia, vaikka, kuten kaikki sammakkoeläimet, he tarvitsivat korkeaa ilmankosteutta ja vesistöjen läheisyyttä kutemiseen. Useiden tutkijoiden mukaan pienistä labyrinttihousuista, jotka pysyivät vesistöjen rannoilla ja pakenivat maavihollisia hyppäämällä veteen, tuli anuran (Anura) esi-isiä, jotka luultavasti syntyivät myöhään permin aikana. Madagaskarin varhaistriaskauden vanhimmalla tunnetulla hännänttömän sammakkoeläimen Triadoba-trachus-edustajalla oli jo sammakoille tyypilliset rungon mittasuhteet ja perusrakenteelliset piirteet, vaikka sillä oli vielä pieni häntä.
Kaksi muuta nykyaikaisten fossiilisten sammakkoeläinten luokkaa tunnetaan paljon myöhemmistä kerrostumista: caudate (Urodela) keskijurakaudelta, jalkattomat tai caecilians (Apoda) varhaisjurakaudelta. Niiden alkuperäongelma on edelleen kiistanalainen. Monien luurangon ja pään lihasten merkkien mukaan pyrstö sammakkoeläimet ovat lähellä anuraaneja ja mahdollisesti niiden kanssa yhteistä alkuperää.
Jalattomat sammakkoeläimet erottuvat hieman toisistaan ja säilyttävät monilta osin alkeellisimman rakenteen nykyaikaisten sammakkoeläinten joukossa. Jotkut paleontologit yhdistävät niiden alkuperän stegokefalien fylogeneettiseen runkoon, jotka yhdistyvät Lepospondyli-alaluokkaan (Lepospondyli). (Toisen näkökulman mukaan kaikilla kolmella nykyaikaisten sammakkoeläinryhmällä oli yhteinen alkuperä, luultavasti labyrinttihousuista).
Lepospondylien joukossa oli monia muotoja, joilla oli voimakkaasti pitkänomainen runko ja heikosti kehittyneet raajat. Tällaisia ovat esimerkiksi lysorofit (Lysorophia). Toisen lepospondyylilajin edustajat - haikarat (Aistopoda), kuten caecilians, menettivät raajansa kokonaan. He eivät kuitenkaan voineet olla caecilian esi-isiä, koska heillä oli useita erityisiä rakenteellisia piirteitä. Nectridia (Nectridia) kuuluu myös lepospondyliciin. Yksi heidän perheistään erottui omituisesta kallon rakenteesta, jonka takakulmat kasvoivat suunnattomasti sivuttain ja taaksepäin pitkien kartiomaisten kasvainten muodossa, joten pää kokonaisuudessaan näytti ylhäältä katsottuna bumerangilta. Oletetaan, että nämä kasvut voisivat toimia eräänlaisena kantosiipialaisina, jotka loivat liikkeen aikana jonkin verran nostoa, mikä edesauttoi liukumista vedessä.
Jo devonikaudella primitiivisistä labyrintodonteista erottui antrakosaurusryhmä (Anthracosauria), jonka vanhin edustaja Tulerpeton löydettiin Tulan alueen ylä-devonin esiintymistä. Aluksi nämä stegokefalaat asuivat myös vesistöjen rannikkoalueella. Antrakosaurusten joukossa suuntaus kohti maanpäällisten elinympäristöjen kehittymistä hahmotettiin kuitenkin hyvin varhain. Kuten useimmissa tapauksissa, tämä suuntaus kehittyi itsenäisesti ja rinnakkain useissa erilaisissa fyleettisissä suvuissa, joista jokainen kehitti vähitellen pitkälti samanlaisia sopeutumisia elämään maalla. Antrakosaurust ja niiden jälkeläiset, jotka maanpäällisestä elämäntavastaan huolimatta säilyttävät yleisen organisoitumisen tason sekä sammakkoeläinten lisääntymiselle ja ontogeneesille ominaiset piirteet (kutevat vesistöissä ja vedessä elävät toukat kidusten kanssa), yhdistetään alaluokkaan batrakosaurukset tai "liskot-sammakkoeläimet" (Batrachosauria). Seymouria (Seymouria; kuva 51), Kot-lassia (Kotlassia) ja siihen liittyvät muodot, jotka olivat olemassa permikaudella, kuuluvat batrakosauruksiin. Kauan aikaa seymourialaisia ei pidetty sammakkoeläiminä, vaan matelijoina, ne ovat kallon, selkärangan ja raajojen rakenteessa niin lähellä todellisia matelijoita. Myöhemmin löydettiin kuitenkin näiden eläinten toukkavaiheen jäänteet, joissa oli jälkiä seismosensoristen elinten kanavista kallon luissa ja jotka siksi elivät vedessä.
Joistakin varhaisen hiilikauden primitiivisistä batrakosauruksista syntyivät todelliset matelijat (Reptilia), joihin kuuluu erityisesti pieni Westlothiana-muoto, joka löydettiin Skotlannin alahiilestä. Sen organisaatio yhdistää kallon progressiiviset (matelijat) piirteet ja useiden selkärangan ja raajojen ominaisuuksien primitiivisen tilan ("mosaiikki"). R. Carroll uskoo, että matelijoiden välittömät esi-isät saattoivat olla pääpiirteiltään lähellä myöhäistä hiilimetsää (Gcphyrostegidae, kuva 51), jotka kuitenkin olivat olemassa liian myöhään ollakseen matelijoiden todellisia esi-isiä. On mahdollista, että Gefirostegit olivat jäänneryhmä, joka säilytti joukon arkaaisia rakenteellisia piirteitä, jotka ovat tyypillisiä matelijoiden varhaishiilikauden esivanhemmille.