Изменение активности почвенных ферментов под действием. Ферментативная активность почв
Цель работы ‑ определение биологической активности почв на разном удалении от дороги по четырем ферментным системам: дегидрогеназам, каталазе, инвертазе, уреазе.
Основные понятия
Почвенно-энзимологические методы позволяют определять не количественное содержание ферментов в почве, а активность ферментов, находящихся преимущественно в адсорбированном (иммобилизованном) состоянии на поверхности почвенных коллоидов и частично в почвенном растворе.
Принципметода определения активности почвенных ферментов основан на учете количества переработанного в процессе реакции субстрата или образующегося продукта реакции в оптимальных условиях температуры, рН среды и концентрации субстратов.
Ферменты, относящиеся к классу оксидоредуктаз, катализируют окислительно-восстановительные реакции, играющие ведущую роль в биохимических процессах в клетках живых организмов, а также в почве. Наиболее распространены в почвах такие оксидо-редуктазы, как каталаза и дегидрогеназы, активность которых является важным показателем генезиса почв.
Каталазаразлагает на воду и молекулярный кислород ядовитую для клетки перекись водорода, образующуюся в процессе дыхания живых организмов в результате различных биохимических реакций окисления органических веществ.
Активность каталазы определяется газометрическим методом по объему выделившегося кислорода, основанным на измерении скорости разложения перекиси водорода при ее взаимодействии с почвой.
Дегидрогеназы ‑ ферменты, которые участвуют в процессе дыхания, отщепляя водород от окисляемых субстратов. Одни дегидрогеназы переносят водород непосредственно на молекулярный кислород, другие - на какие-либо акцепторы, например на хиноны, метиленовую синь.
Для определения активности дегидрогеназы в качестве акцептора водорода применяют бесцветные соли тетразолия (2,3,5-трифенилтетразолий хлористый (ТТХ), которые восстанавливаются в красные соединения формазана (трифенилформазан (ТФФ).
Гидролазы осуществляют реакции гидролиза разнообразных сложных органических соединений, действуя на различные связи: сложноэфирные, глюкозидные амидные, пептидные и др. К этому классу относятся ферменты инвертаза, уреаза и др., активность которых является важным показателем биологической активности почв и широко используется для оценки антропогенного воздействия.
Инвертаза действует на p-фруктофуранозидную связь в сахарозе, рафинозе, стахиоэе и производит расщепление сахарозы на эквимолярные количества глюкозы и фруктозы.
Фотоколориметрическое определение активности инвертазы основано на учете восстанавливающих сахаров, образующихся при расщеплении сахарозы.
Разложение органических азотистых соединений осуществляется при непосредственном участии внеклеточных ферментов. Образующийся при уреазной активности аммиак служит источником питания растений.
Уреаза катализирует гидролиз мочевины. Конечными продуктами гидролиза являются аммиак и углекислый газ. Мочевина попадает в почву в составе растительных остатков, навоза и как азотное удобрение; она образуется также в самой почве в качестве промежуточного продукта в процессе превращения азотистых органических соединений - белков и нуклеиновых кислот.
Определение каталазной активности
Оборудование и реактивы:
Cистема для газометрии (рис. 8); 10%-й раствор Н 2 O 2 ; СаСО э.
Рис. 8 ‑ Установка для газометрического определения каталазной активности в почвенных образцах:
1 - колба, 2 - бюретка, 3 - переходник, 4 - груша с водой
Порядок выполнения работы
1. Навеску просеянной почвы 1 г внести в колбу на 100 см 3 , добавить 0,5 г СаСО 3 .
2. На дно осторожно поставить с помощью пинцета маленький стаканчик с 1,7 см 3 10%-го раствора перекиси водорода.
3. Навеску почвы смочить 4 см 3 дистиллированной воды.
4. Колбу плотно закрыть каучуковой пробкой с трубкой, соединенной с бюреткой толстостенным каучуком через тройник, снабженный зажимом. Бюретка сообщается с грушей. Бюретка и груша заполнены водой. Уровень воды в них уравновешивают и грушу закрепляют на определенной высоте.
5. Начало опыта отметить по секундомеру в момент, когда сосудик с перекисью водорода опрокинут, и вслед за этим встряхнуть содержимое колбы. Взбалтывание смеси следует продолжать во все время опыта, не касаясь непосредственно дна колбы руками. Выделяющийся кислород вытесняет из бюретки воду, уровень которой отмечают.
6. Количество выделившегося молекулярного кислорода учитывают в течение 1 мин при температуре 18-20 0 С.
7. Активность каталазы выражают в объеме (см 3) кислорода, выделившегося на 1 г почвы в минуту. Ошибка определения до 5%.
8. Аналогичные процедуры проделать со всеми образцами почв.
9. По табл. 15 оценить степень насыщения исследуемых почв каталазой.
Таблица15 ‑ Шкала для оценки степени обогащенности почв ферментами
| Степень обогашенности почв | Каталаза, О 2 см 3 /г за 1 мин | Дегидрогеназы, мг ТФФ на 10 г за 24 ч | Инвертаза, мг глюкозы на 1 г за 24 ч | Уреаза, мг NH 4 , на 10 г за 24 ч | Фосфотаза, мг Р 2 О 3 на 10 г за 1 ч |
| Очень бедная | < 1 | <1 | <5 | <3 | <0,5 |
| Бедная | 1-3 | 1-3 | 5-15 | 3-10 | 0,5-1,5 |
| Средняя | 3-10 | 3-10 | 15-50 | 10-30 | 1,5-5,0 |
| Богатая | 10-30 | 10-30 | 50-150 | 30-100 | 5-15 |
| Очень богатая | >30 | >30 | > 150 | > 100 | > 15 |
Определение дегидрогиназной активности
Приборы, посуда, реактивы :
Фотоколориметр; миллиметровая бумага; 0,1М раствор глюкозы; 1 %-й раствор 2,3,5-трифенилтетразолия хлористого (ТТХ); СаСО 3 ; этиловый спирт; трифенилформазан (ТФФ).
Порядок выполнения работы
1. Навески воздушно-сухой почвы по 1 г из каждого образца поместить в пробирки, добавить по 10 мг (на кончике шпателя) СаСО 3 , по 1 см 3 0,1 М раствора глюкозы и по 1 см 3 1%-го раствора ТТХ; содержимое каждой пробирки тщательно смешать.
2. Пробирки поместить в анаэростат и откачать воздух насосом при разрежении 10-12 мм рт. ст. в течение 2-3 мин. Затем инкубировать при 30 0 С в течение 24 ч.
3. По истечении времени инкубации содержимое пробирок экстрагировать в 3-4 приема 25 см 3 этилового спирта. Для этого небольшой объем спирта внести в пробирку и встряхивать в течение 5 мин до появления красной окраски. Дать отстояться и надпочвенную жидкость профильтровать через бумажный фильтр. Добавить в пробирку следующую порцию спирта.
4. Полученный окрашенный раствор формазана колориметрировать на ФЭКе с синим светофильтром (500-600 нм).
5. Количество формазана в миллиграммах рассчитать по стандартной кривой. Для этого приготовить стандартный раствор формазана в этиловом спирте в концентрации 0,1 мг в 1 см 3 . Рабочие растворы для составления кривой приготовить путем разведений стандартного раствора (примерно 5 точек). Стандартную кривую построить на миллиметровой бумаге в системе: оптическая плотность при длине волны 500-600 нм - концентрация формазана в спирте.
6. Вычислить активность дегидрогеназы. По табл. 15 оценить степень насыщения исследуемых почв дегидрогеназой.
Обработка данных
Активность дегидрогеназы (X) выражают в миллиграммах ТФФ на 10 г почвы за сутки по формуле:
где V ‑ общий объем фильтрата, 25 см 3 ;
10 ‑ пересчетный коэффициент веса почвы, г;
v ‑ произведение объемов субстрата и реагента, 1 см 3 ;
А ‑ количество ТФФ, полученное по калибровочной кривой, мг/см 3 . Ошибка определения ‑ до 8 %.
Определение инвертазной активности
Приборы, посуда, реактивы :
Фотоколориметр; 5%-й раствор сахарозы; ацетатный буфер (рН 4,7); толуол; раствор Феллинга: а ‑ 40 г CuSO 4 ×5Н 2 О растворяют в воде и доводят до 1 дм 3 , фильтруют через бумажный фильтр, б ‑ 200 г сегнетовой соли (С 4 H 4 O 6 KNa×4Н 2 О) растворяют в дистиллированной воде, прибавляют 150 г КОН и доводят до 1 дм 3
Порядок выполнения работы
1. В колбы вместимостью 50 см 3 поместить по 5 г каждого образца почвы, добавить по 10 см 3 5%-го раствора сахарозы, 10 мл ацетатного буфера (рН 4,7) и 5-6 капель толуола.
2. Колбы закрыть пробками, встряхнуть, поместить в термостат при температуре 30 0 С на 24 ч и периодически встряхивать их.
3. После инкубации содержимое колб отфильтровать в мерные колбы на 25 см 3 . Довести до метки.
4. Из фильтратов взять по 6 см 3 в большие пробирки, добавить по 3 см 3 раствора сегнетовой соли и 3 см 3 раствора сернокислой меди, хорошо перемешать и кипятить на водяной бане 10 мин. Получается красный осадок.
5. Пробирки с раствором охладить в воде, содержимое отфильтровать в большие пробирки. Прозрачный фильтрат колориметрировать на ФЭК, используя светофильтр с длиной волны 630 нм, ширина кюветы 1 см.
6. Для получения калибровочной кривой приготовить стандартный раствор: 6 мг глюкозы в 1 см 3 . Разведением приготовить серию растворов. Фотоколориметрировать и построить кривую: оптическая плотность ‑ концентрация глюкозы в 1 см 3 .
7. Вычислить активность и по табл. 15 оценить степень насыщения исследуемых почв инвертазой.
Обработка данных
Активность инвертазы (X) выражают в миллиграммах глюкозы на 1 г почвы за 24 ч по формуле:
где А ‑ количество глюкозы, полученное по калибровочной кривой из оптической плотности, мг/см 3 ;
m ‑ навеска почвы, 5 г;
V ‑ общий объем фильтрата, 25 см 3 ;
v ‑ объем фильтрата, взятого для анализа, 6 см 3 .
Ошибка определения ‑ до 5 %.
Определение уреазной активности почв
Приборы, посуда, реактивы :
Фотоколориметр; 2%-й раствор мочевины в фосфатном буфере (рН = 6,7); 50%-й раствор сегнетовой соли; 50%-й раствор CCl 3 COOH (трихлоруксусная кислота); 1%-й раствор КС1; реактив Несслера; стандартный раствор NH 4 C1.
Порядок выполнения работы
1. По 5 г воздушно-сухой почвы поместить в колбы емкостью 100 см 3 , прилить по 20 см 3 2%-го раствора мочевины в фосфатном буфере (рН 6,7) и по 200 мкл толуола.
2. Колбы плотно закрыть и поместить в термостат при температуре 37 0 С на 4 ч.
3. После экспозиции прилить по 1 см 3 50%-го раствора трихлоруксусной кислоты.
4. Для вытеснения из почвы поглощенного аммиака добавить по 50 см 3 1 н. раствора хлористого калия.
5. Содержимое колб отфильтровать.
6. По 2 см 3 фильтрата поместить в мерные колбы объемом 50 см 3 , развести водой до 30 см 3 , затем прилить по 2 см 3 50%-го раствора сегнетовой соли и по 2 см 3 реактива Несслера. Колбы долить водой до метки, перемешать и окрашенный раствор колориметрировать при длине волны 400 нм.
8. Вычисляют активность уреазы.
9. По табл. 15 оценить степень насыщения исследуемых почв уреазой.
Обработка данных
Активность уреазы (X) выражают в миллиграммах N-NH 4 на 1 г почвы за 4 ч по формуле:
V ‑ общий объем фильтрата, 50 см 3 ;
m - навеска почвы, 5 г.
Вопросы для самоподготовки:
1. Что такое каталазная активность?
2. Дайте оределение инвертазной активности.
3. Охарактеризуйте уреазную активность.
4. Что такое буферная смесь?
5. Принцип и сущность метода определения активности почвенных ферментов.
6. Методика отбора образцов почвы.
ПРИЛОЖЕНИЯ
Таблица 1 ‑ Примерный список организмов - индикаторов сапробности
| Организмы | Сапробность |
| Нитчатые бактерии: | |
| Sphaerotilus natans | р |
| Beggiatoa sp. | р |
| Thiothrix sp. | р |
| Грибы: | |
| Leptomitus lacteus | α |
| Mucor racemosus | α |
| Fusarium aquaeductum | р |
| Водоросли: | |
| сине-зеленые: | |
| Anabaena flos aquae | β |
| Microcystis aeruginosa | β |
| Aphanizomenon flos aquae | β |
| Oscillatorla tenuis | α |
| Диатомовые | - |
| Cymbella cesati | о |
| Oomphonema cevli | о |
| Melostra granulata | β |
| Navicula angustata | α |
| Navicula apiculata | α |
| Synedra acus | β |
| Synedra ulna | β |
| Nitzschia palea | α |
| эвгленовые: | |
| Euglena acus | β |
| Euglena viridis | р |
| Euglena deses | α |
| зеленые и протококковые: | |
| Volvox globator | о-β |
| Ankistrodesmus falcatus | β-α |
| Crucigenta rectangularis | а-β |
| Scenedesmus quadricauda | β |
| Draparnaldia sp. | о |
| Ulothrix zonata | о |
| Stlgeoclonium tenue | α |
| Животные: | |
| амебы: | |
| Pelornyxa palustris | р |
| Организмы | Сапробность |
| инфузории: | |
| Colpidium, campylum | p |
| Colpldlum colpoda | p |
| Euplotes charon | β |
| Chllodon cucullulus | p |
| Opercularia coaretata | α |
| Paramecium caudatum | α |
| Spirostomum amblguum | α |
| Stentor coeruleus | α |
| Vortlcella convallarla | α |
| Vorticella microstoma | p |
| Podophrya fixa | α |
| коловратки: | |
| Kellcottia longispina (syn. Notholca Iongispina) | о |
| Keratella cochlearls | β |
| Keratella quadrata | β |
| Leucane lunarls (syn. Monostyla lunarls) | β |
| Rotaria rotatoria (syn. Rotifer vulgaris) | α |
| олигохеты: | |
| Limnodrilus hofmelsterl | p |
| Tub if ex tublfex | p |
| Stylarla lacustris | β |
| ракообразные: | |
| Daplmla magna | α |
| Daphnla pulex | α |
| Leptodora Kindtli | о |
| Eudiaptomus gracilis | o |
| Astacus fluviatilis | o |
| насекомые: | |
| Caenls macrura | o |
| Heptagenia coerulana | β |
| Chironomus Plumosus | р |
| рыбы: | |
| лещ: | β |
| усач | β |
| форель | o |
| линь | β-α |
Таблица 2 ‑ Шкала частот для пересчета организмов в 100 полях на частоту
| Значение частоты | Микробентос | Обрастания | |
| Данные подсчета | Сумма в 100 полях | ||
| 1-я категория крупности | |||
| Не более 1 в каждом 2-м поле зрения Не более 2 в поле зрения Не более 10 в поле зрения Не более 30 в поле зрения Не более 60 в поле зрения Более 60 в поле зрения | Не более 1 в каждом 2-м поле зрения Не более 2 в поле зрения Не более 10 в поле зрения Не более 50 в поле зрения Не более 250 в поле зрения Более 250 в поле зрения | 1-50 50-200 200-1000 1000-5000 5000-25000 Более 25000 | |
| 2-я категория крупности | |||
| Не более 1 в каждом 20-м поле зрения Не более 1 в каждом 5-м поле зрения Не более 1 в поле зрения Не более 3 в поле зрения Не более 6 в поле зрения Более 6 в поле зрения | Не более 2 в 20 полях зрения Не более 1 в 5 поле зрения Не более 1 в поле зрения Не более 5 в поле зрения Не более 25 в поле зрения Более 25 в поле зрения | 1-5 6-20 21-100 100-500 500-2500 Более 2500 | |
| 3-я категория крупности | |||
| 1 в 100 полях зрения 1 в 50 полях зрения Не более 1 в 10 полях зрения Не более 1 в 4 полях зрения Не более 1 в 2 полях зрения Приблизительно 1 в поле зрения | 1 в 100 полях зрения 1 в 50 полях зрения Не более 1 в 10 в полях зрения 1в2 полях зрения Не более 2 в поле зрения Более 2 в поле зрения | 3-10 10-50 50-200 Более 200 |
Приложение
Таблица 13. Пересчет результатов количественного учета на значение частоты
Приложение
Пример вычисления сапробности
Проба: река ниже города. Дата ________________ Сообщество: обрастания.
| Организмы | s | h | sft |
| Euglena viridis | p | ||
| Scenedesmus acuminatus | β | ||
| Spirogyra sygmoidea | β | ||
| Closterium acerosum | α | ||
| Closterium moniliierum | β | ||
| Cyclotella menengiana | α | ||
| Cymbella vesiculosa | β | ||
| Diatoma vulgare | β | ||
| Melosira italica | β | ||
| Melosira varians | β | ||
| Navicula cryptocephala | α | ||
| Navicula viridua | α | ||
| Nitzschia acicularis | β | ||
| Nitzschia palea | α | ||
| Surirella ovata | β | ||
| Chilidonella cuculata | α | ||
| Colpoda cuculus | α | ||
| Sh=41 | S(sh)=103 |
Sh p =3; Sh α =15; Sh β =23.
S=S(sh)/(Sh)-103/41=2,51/
Вычисление погрешности:
Интервал точности для статистической надежности 95%.
S=s±t 0,05 s S =2,51±2,02×0,1;
Похожая информация.
Ферменты - это катализаторы химических реакций белковой природы, отличающиеся специфичностью действия в отношении катализа определенных химических реакций. Они являются продуктами биосинтеза всех живых почвенных организмов: древесных и травянистых растений, мхов, лишайников, водорослей, микроорганизмов, простейших, насекомых, беспозвоночных и позвоночных животных, представленных в природной обстановке определенными совокупностями - биоценозами.
Биосинтез ферментов в живых организмах осуществляется благодаря генетическим факторам, ответственным за наследственную передачу типа обмена веществ и его приспособительную изменчивость. Ферменты являются тем рабочим аппаратом, при помощи которого реализуется действие генов. Они катализируют в организмах тысячи химических реакций, из которых в итоге слагается клеточный обмен. Благодаря им химические реакции в организме осуществляются с большой скоростью.
В настоящее время известно более 900 ферментов. Их подразделяют на шесть главных классов.
1. Оксиредуктазы, катализирующие окислительно-восстановительные реакции.
2. Трансферазы, катализирующие реакции межмолекулярного переноса различных химических групп и остатков.
3. Гидролазы, катализирующие реакции гидролитического расщепления внутримолекулярных связей.
4. Лиазы, катализирующие реакции присоединения групп по двойным связям и обратные реакции отрыва таких групп.
5. Изомеразы, катализирующие реакции изомеризации.
6. Лигазы, катализирующие химические реакции с образованием связей за счет АТФ (аденозинтрифосфорной кислоты).
При отмирании и перегнивании живых организмов часть их ферментов разрушается, а часть, попадая в почву, сохраняет свою активность и катализирует многие почвенные химические реакции, участвуя в процессах почвообразования и в формировании качественного признака почв - плодородия. В разных типах почв под определенными биоценозами сформировались свои ферментативные комплексы, отличающиеся активностью биокаталитических реакций.
В. Ф. Купревич и Т. А. Щербакова (1966) отмечают, что важной чертой ферментативных комплексов почв является упорядоченность действия имеющихся групп ферментов, которая проявляется в том, что обеспечивается одновременное действие ряда ферментов, представляющих различные группы; исключаются образование и накопление соединений, имеющихся в почве в избытке; излишки накопившихся подвижных простых соединений (например, NH 3) тем или иным путем временно связываются и направляются в циклы, завершающиеся образованием более или менее сложных соединений. Ферментативные комплексы являются уравновешенными саморегулирующимися системами. В этом основную роль играют микроорганизмы и растения, постоянно пополняющие почвенные ферменты, так как многие из них являются короткоживущими. О количестве ферментов косвенно судят по их активности во времени, которая зависит от химической природы реагирующих веществ (субстрата, фермента) и от условий взаимодействия (концентрации компонентов, рН, температуры, состава среды, действия активаторов, ингибиторов и т.д.).
В данной главе рассматривается участие в некоторых химических почвенных процессах ферментов из класса гидролаз - активность инвертазы, уреазы, фосфатазы, протеазы и из класса оксиредуктаз - активность каталазы, пероксидазы и полифенолоксидазы, имеющих большое значение в превращении азот- и фосфорсодержащих органических веществ, веществ углеводного характера и в процессах образования гумуса. Активность этих ферментов - существенный показатель плодородия почв. Кроме того, будет охарактеризована активность этих ферментов в лесных и пахотных почвах разной степени окультуренности на примере дерново-подзолистых, серых лесных и дерново-карбонатных почв.
ХАРАКТЕРИСТИКА ПОЧВЕННЫХ ФЕРМЕНТОВ
Инвертаза - катализирует реакции гидролитического расщепления сахарозы на эквимолярные количества глюкозы и фруктозы, воздействует также на другие углеводы с образованием молекул фруктозы - энергетического продукта для жизнедеятельности микроорганизмов, катализирует фруктозотрансферазные реакции. Исследования многих авторов показали, что активность инвертазы лучше других ферментов отражает уровень плодородия и биологической активности почв.
Уреаза- катализирует реакции гидролитического расщепления мочевины на аммиак и диоксид углерода. В связи с использованием мочевины в агрономической практике необходимо иметь в виду, что активность уреазы выше у более плодородных почв. Она повышается во всех почвах в периоды их наибольшей биологической активности - в июле - августе.
Фосфатаза (щелочная и кислая) - катализирует гидролиз ряда фосфорорганических соединений с образованием ортофосфата. Активность фосфатазы находится в обратной зависимости от обеспеченности растений подвижным фосфором, поэтому она может быть использована как дополнительный показатель при установлении потребности внесения в почвы фосфорных удобрений. Наиболее высокая фосфатазная активность в ризосфере растений.
Протеазы - это группа ферментов, при участии которых белки расщепляются до полипептидов и аминокислот, далее они подвергаются гидролизу до аммиака, диоксида углерода и воды. В связи с этим протеазы имеют важнейшее значение в жизни почвы, так как с ними связаны изменение состава органических компонентов и динамика усвояемых для растений форм азота.
Каталаза - в результате ее активирующего действия происходит расщепление перекиси водорода, токсичной для живых организмов, на воду и свободный кислород. Большое влияние на каталазную активность минеральных почв оказывает растительность. Как правило, почвы, находящиеся под растениями с мощной глубоко проникающей корневой системой, характеризуются высокой каталазной активностью. Особенность активности каталазы заключается в том, что вниз по профилю она мало изменяется, имеет обратную зависимость от влажности почв и прямую - от температуры.
Полифенолоксидаза и пероксидаза - им в почвах принадлежит важная роль в процессах гумусообразования. Полифенолоксидаза катализирует окисление полифенолов в хиноны в присутствии свободного кислорода воздуха. Пероксидаза же катализирует окисление полифенолов в присутствии перекиси водорода или органических перекисей. При этом ее роль состоит в активировании перекисей, поскольку они обладают слабым окисляющим действием на фенолы. Далее может происходить конденсация хинонов с аминокислотами и пептидами с образованием первичной молекулы гуминовой кислоты, которая в дальнейшем способна усложняться за счет повторных конденсаций (Кононова, 1963).
Замечено (Чундерова, 1970), что отношение активности полифенолоксидазы (S) к активности пероксидазы (D), выраженное в процентах (), имеет связь с накоплением в почвах гумуса, поэтому эта величина получила название условный коэффициент накопления гумуса (К). У пахотных слабоокультуренных почв Удмуртии за период с мая по сентябрь он составил: у дерново-подзолистой - 24 %, у серой лесной оподзоленной - 26 и у дерново-карбонатной почвы - 29 %.
ФЕРМЕНТАТИВНЫЕ ПРОЦЕССЫ В ПОЧВАХ
Биокаталитическая активность почв находится в значительном соответствии со степенью обогащенности их микроорганизмами (табл. 11), зависит от типа почв и изменяется по генетическим горизонтам, что связано с особенностями изменения содержания гумуса, реакции, Red-Ox-потенциала и других показателей по профилю.
В целинных лесных почвах интенсивность ферментативных реакций в основном определяют горизонты лесной подстилки, а в пахотных - пахотные слои. Как в одних, так и в других почвах все биологически менее активные генетические горизонты, находящиеся под горизонтами А или А п, имеют низкую активность ферментов, незначительно изменяющуюся в положительную сторону при окультуривании почв. После освоения лесных почв под пашню ферментативная активность образованного пахотного горизонта по сравнению с лесной подстилкой оказывается резко сниженной, но по мере его окультуривания повышается и в сильно окультуренных видах приближается или превышает показатели лесной подстилки.
11. Сопоставление биогенносга и ферментативной активности почв Среднего Предуралья (Пухидская, Ковриго, 1974)
|
№ разреза, название почвы |
Горизонт, глубина взятия образца, см |
Общее количество микроорганизмов, тыс. на 1 г абс. сух. почвы (в среднем за 1962, 1964-1965 гг.) |
Показатели активности ферментов (в среднем за 1969-1971 гг.) |
|||||||
|
Инвертаза, мг глюкозы на 1 г почвы за I сут |
Фосфатаза, мг фенолфталеина на 100 г почвы за 1 ч |
Уреаза, мг NH, нa 1 г почвы за 1 сут |
Каталаза, мл 0 2 на 1 г почвы за 1 мин |
Полифенолоксидаза |
Пероксидаза |
|||||
|
мг пурпурогаллина на 100 г почвы |
||||||||||
|
3. Дерново-среднеподзолистая среднесуглинистая (под лесом) |
||||||||||
|
Не определяли |
||||||||||
|
1.Дерново-средне-подзолистая средне-суглинистая слабоокультуренная |
||||||||||
|
10.Сераялесная оподзоленная тяжел осуглинистая слабоокультуренная |
||||||||||
|
2. Дерново-карбонатная слабовыщело-ченная л егкосуглинистая слабоокультуренная |
||||||||||
Активность биокаталитических реакций почв изменяется. Наименьшая она весной и осенью, а наиболее высокая обычно в июле-августе, что соответствует динамике общего хода биологических процессов в почвах. Однако в зависимости от типа почв и их географического положения динамика ферментативных процессов весьма различна.
Контрольные вопросы и задания
1. Какие соединения называют ферментами? Каковы их продуцирование и значение для живых организмов? 2. Назовите источники почвенных ферментов. Какую роль играют отдельные ферменты в почвенных химических процессах? 3. Дайте понятие о ферментативном комплексе почв и его функционировании. 4. Дайте общую характеристику течения ферментативных процессов в целинных и пахотных почвах.
Из многочисленных показателей биологической активности почвы большое значение имеют почвенные ферменты. Их разнообразие и богатство делают возможным осуществление последовательных биохимических превращений, поступающих в почву органических остатков.
Название «фермент» происходит от латинского «ферментум» – брожу, закваска. Явление катализа и в настоящее время полностью не разгадано. Сущность действия катализатора заключается в снижении энергии активации, необходимой для химической реакции, направляя ее обходным путем через промежуточные реакции, которые требуют меньшей энергии, идущие без катализатора. Благодаря этому повышается и скорость основной реакции. Под действием фермента ослабляются внутримолекулярные связи в субстрате вследствие некоторой деформации его молекулы, происходящей при образовании промежуточного комплекса фермент-субстрата.
Таким образом, роль ферментов заключается в том, что они значительно ускоряют биохимические реакции и делают их возможными при обычной нормальной температуре.
Ферменты, в отличие от неорганических катализаторов, обладают избирательностью действия. Специфичность действия ферментов выражается в том, что каждый фермент действует лишь на определенное вещество, или же на определенный тип химической связи в молекуле. По своей биохимической природе все ферменты – высокомолекулярные белковые вещества. На специфичность ферментных белков влияет порядок чередования в них аминокислот. Некоторые ферменты помимо белка содержат более простые соединения. Например, в составе различных окислительных ферментов содержатся органические соединения железа. В состав других входят медь, цинк, марганец, ванадий, хром, витамины и другие органические соединения.
В основу единой классификации ферментов положена специфичность к типу реакции, и в настоящее время ферменты подразделяют на 6 классов. В почвах наиболее изучены оксидоредуктазы (катализируют процессы биологического окисления) и гидролазы (катализируют расщепление с присоединением воды). Из оксидоредуктаз в почве наиболее распространены катал аза, дегидрогеназы, фенолоксидазы и др.
Они участвуют в окислительно-восстановительных процессах синтеза гумусовых компонентов. Из гидролаз наиболее широко в почвах распространены инвертаза, уреаза, протеаза, фосфатазы. Эти ферменты участвуют в реакциях гидролитического распада высокомолекулярных органических соединений и тем самым играют важную роль в обогащении почвы подвижными и доступными растениям и микроорганизмам питательными веществами.Исследованием ферментативной активности почв занималось большое количество исследователей. В результате исследований доказано, что ферментативная активность – это элементарная почвенная характеристика. Ферментативная активность почвы складывается в результате совокупности процессов поступления, иммобилизации и действия ферментов в почве. Источниками почвенных ферментов служит все живое вещество почв: растения, микроорганизмы, животные, грибы, водоросли и т. д. Накапливаясь в почве, ферменты становятся неотъемлемым реактивным компонентом экосистемы. Почва является самой богатой системой по ферментному разнообразию и ферментативному пулу. Разнообразие и богатство ферментов в почве позволяет осуществляться последовательным биохимическим превращениям различных поступающих органических остатков.
Значительную роль почвенные ферменты играют в процессах гумусообразования. Превращение растительных и животных остатков в гумусовые вещества является сложным биохимическим процессом с участием различных групп микроорганизмов, а также иммобилизованных почвой внеклеточных ферментов. Выявлена прямая связь между интенсивностью гумификации и ферментативной активностью.
Особо следует отметить значение ферментов в тех случаях, когда в почве складываются экстремальные для жизнедеятельности микроорганизмов условия, в частности при химическом загрязнении. В этих случаях метаболизм в почве остается в известной мере неизменным благодаря действию иммобилизированных почвой, и поэтому устойчивых, ферментов. Максимальная каталитическая активность отдельных ферментов наблюдается в относительно небольшом интервале pH, который является для них оптимальным. Поскольку в природе встречаются почвы с широким диапазоном реакции среды (pH 3,5-11,0), то их уровень активности весьма различен.
Исследованиями различных авторов установлено, что активность почвенных ферментов может служить дополнительным диагностическим показателем почвенного плодородия и его изменения в результате антропогенного воздействия. Применению ферментативной активности в качестве диагностического показателя способствуют низкая ошибка опытов и высокая устойчивость ферментов при хранении образцов.
Инвертаза - катализирует реакции гидролитического расщепления сахарозы на эквимолярные количества глюкозы и фруктозы, воздействует также на другие углеводы с образованием молекул фруктозы - энергетического продукта для жизнедеятельности микроорганизмов, катализирует фруктозотрансферазные реакции. Исследования многих авторов показали, что активность инвертазы лучше других ферментов отражает уровень плодородия и биологической активности почв.[ ...]
Анализы инвертазы после 1 года свидетельствуют о дальнейшем уменьшении ее во всех образцах в 2-3 раза в зависимости от типа почв, что, по-видимому, объясняется истощением почвы углеродсодержащими соединениями.[ ...]
Из класса гидролаз изучена активность инвертазы, гидролизирующей сахарозу на глюкозу и фруктозу, и уреазы, катализирующей гидролиз мочевины. Активность этих ферментов в грунте очень низкая, но при внесении торфа увеличивается пропорционально его дозам и мало зависит от количества минеральных удобрений. Следует отметить, что внесение самой большой дозы (ЫРКЦ, а также СаСОэ не имеет преимуществ перед меньшими дозами удобрений в стимулировании активности как гидролаз, так и оксидоредуктаз.[ ...]
Для трассы аэропорт - пос. Кангалассы обратная зависимость между активностью уреазы, инвертазы и протеазы и содержанием свинца не обнаружена. Это свидетельствует об отсутствии ингибирующего эффекта свинца в дозе, не превышающей ПДК. Отмечается параллельное увеличение активности всех ферментов и свинца по мере удаления от источника загрязнения, что в данном случае объясняется увеличением гумусированности почв. Известно, что почвы с высоким содержанием гумуса в большей степени накапливают ТМ и характеризуются повышенной ФА.[ ...]
Соединения данной группы задерживают рост новых побегов, временно снижают активность инвертазы в сахарной свекле и подавляют биосинтез хлорофилла. И все же их первичное действие - это подавление биосинтеза ароматических аминокислот. Соединения типа Ы-фосфонметилглицина подавляют этот синтез, действуя на участках преобразования дегидрохинной и префеновой кислот.[ ...]
По-видимому, образование сахарозы происходит в паренхимных клетках флоэмы, откуда она поступает в ситовидные трубки, которые лишены ферментов, разлагающих сахарозу (инвертазы), что и определяет сохранность этого соединения на всем пути его транспорта.[ ...]
Проведенная работа позволяет заключить, что накопление подвижных форм свинца и никеля в дозах, превышающих ПДК, приводит к снижению активности ферментов в почвах. Понижение активности протеазы, уреазы и инвертазы в почвах обусловливает соответствующее торможение процессов гидролиза белков, мочевины и олигосахаридов, что в целом приводит к снижению биологической активности почв. Изменение ФА -перспективный метод диагностики экологического состояния почв. Из рассмотренных нами ферментов наиболее высокие диагностические свойства проявляет уреаза.[ ...]
Состояние почв оценивалось двумя биоиндикационными методами: по ферментативной активности почв и мутационному воздействию почв на тест-объект. В городских почвогрунтах определялась активность трех ферментов - инвертазы, каталазы и уреазы (Хазиев, 1990), из которых наиболее вариабильной оказалась активность уреазы. По этой причине для интегральной оценки выбраны показатели именно этого фермента, активность которого в значительной степени зависела от концентрации в почве широкого круга поллютантов.[ ...]
Гистохимические анализы позволили установить общность окислительного режима пыльцы и пыльцевых трубок у различных представителей покрытосеменных растений. При этом установлено, что наиболее интенсивно биохимические процессы протекают в кончике пыльцевой трубки.[ ...]
Другая группа эвокациопыых изменений связана с активацией энергетических процессов, необходимых для реализаций морфогенетической программы репродуктивного развития.[ ...]
При внесении больших норм ГХБД и в жидкой, и в гранулированной форме угнетение развития отдельных групп микроорганизмов не проходит и к полутора годам, прошедшим после фумигации. Активность почвенных ферментов (каталазы и инвертазы) к этому времени составляет по этим (вариантам опыта 70-80% от активности ферментов в контрольном варианте. Через 5 месяцев после внесения больших норм ГХБД (жидкого и гранулированного) снижается содержание в почве нитратов, что свидетельствует об угнетении процесса нитрификации.[ ...]
Агрохимические свойства почв определяли общепринятыми методами , pH водной и солевой вытяжек - потенциометрическим, содержание углерода - методом Тюрина, подвижного азота - по Башкину и Кудеярову, подвижного фосфора - по Чирикову, ферментативную активность почв (инвертазу, уреазу и каталазу) - по Хазиеву .[ ...]
У многих представителей лучистых грибков выявлен фермент амилаза, при помощи которого организмы расщепляют крахмал с различной интенсивностью, в зависимости от вида культуры. Одни культуры разлагают крахмал до декстринов, другие - до сахаров. У некоторых актиномицетов обнаружен фермент инвертаза, который расщепляет сахарозу на легкоусвояемые сахара - глюкозу и фруктозу. Отмечено, что проактиномицеты могут усваивать сахарозу без ее разложения.[ ...]
Такие уровни загрязнения отразились и на содержании подвижных, доступных растениям форм соединений тяжелых металлов. Их количество также увеличилось в 1,5-2 и даже в 5 раз. Эти изменения отразились на почвенной биоте, общих свойствах почв и почвенном плодородии. В частности, резко снизилась активность почвенных ферментов: инвертазы, фосфатазы, уреазы, каталазы; примерно в 2 раза снизилось продуцирование С02. Ферментативная активность - хороший интегральный показатель экологической обстановки в системе «почва - растение». На загрязненных почвах резко снизилась и урожайность различных культур. Так, урожай томатов (ц/га) в среднем снизился от 118,4 до 67,2; огурцов - от 68,3 до 34,2; капусты - от 445,7 до 209,0; картофеля - от 151,8 до 101,3; яблок - от 72,4 до 32,6 и персиков - от 123,6 до 60,6.[ ...]
Среди тундровых почв поймы потенциал биохимической активности возрастает от почв прирусловой поймы к центральной и притеррасной. В свою очередь, ферментативная активность в органогенных пойменных почвах выше, чем в минеральных. В гумусовых горизонтах (0-13 см) изученных почв отмечается довольно высокая активность уреазы, инвертазы, фосфатазы и дегидрогеназы - ферментов, участвующих в обменных процессах азота, углеводов, фосфора и окислительно-восстановительных.[ ...]
Активность фосфатазы низкая, а в большинстве случаев фосфатазная активность отсутствует, что связано с очень низким содержанием подвижного фосфора на фоне относительно высокого содержания в перегнойно-торфянистых горизонтах его валовых форм. В отличие от ферментов, участвующих в обменных процессах азота и фосфора, ферменты углеводородного обмена (инвертаза) проявляют свою активность до надмерзлотных горизонтов, что определяется гуму-сированностью профиля.[ ...]
Изменение ферментативной активности почв за четыре года проведения опыта показано в табл. 6.8. Как видно из полученных результатов, активность уреазы и фосфатазы снизилась, но основные закономерности - более высокая активность в вариантах без применения ППС при внесении торфа и минеральных удобрений и отсутствие ферментативной активности в контрольных вариантах - сохраняются. В то же время активность инвертазы, играющей важную роль в круговороте углерода в биогеоценозе, возрастает на четвертый год почти по всем вариантам опыта, в том числе и при внесении ППС, что подтверждает также интенсивность мине-рализационных процессов торфа и универсинов.[ ...]
Очень перспективным методом очистки воды от всевозможных загрязняющих ее веществ, особенно синтетических, является использование иммобилизованных (закрепленных, нерастворимых) ферментов - «ферментов второго поколения». Идея закрепления ферментов на нерастворимом в воде носителе и применения таких мощных катализаторов в технологических процессах и медицине возникла давно. Еще в 1916 г. осуществлена адсорбция инвертазы на активированном угле в свежевыделенной гидроокиси алюминия. С 1951 г. для фракционирования антител и выделения антигенов используют конъюгацию белков с целлюлозой. До недавнего времени существовал единственный метод закрепления ферментов - обыкновенная физическая адсорбция. Однако адсорбционная емкость известных материалов относительно белков явно недостаточна, а силы адгезии невелики, и разрыв связи между ферментом и поверхностью адсорбента может наступать от малейших изменений условий процесса. Поэтому такой метод иммобилизации не нашел широкого применения, но, поскольку он прост и может, по-видимому, способствовать выяснению механизма действия ферментов в живых системах, илах и почве, а в некоторых случаях применяться на практике, некоторые исследователи занимаются изучением адсорбции ферментов, поиском новых, эффективных носителей и т. д. .[ ...]
Если учесть выраженные и длительные физиологические изменения процессов роста и развития, вызываемые этиленом, не покажется удивительным, что происходят также изменения в синтезе РНК и белка и в активности ферментов. Неоднократно проверялась возможность прямого воздействия этилена на активность различных ферментов, например глюкозидазы, а-амилазы, инвертазы и перок-сидазы, но были получены отрицательные результаты-Вместе с тем синтез целого ряда ферментов четко возрастает. К числу ферментов, относительно быстро синтезируемых после воздействия этилена, относится перокси-даза. В плодах цитрусовых усиливается синтез фенил-аланин-аммиак-лиазы, причем С02 и ингибиторы транскрипции блокируют этот процесс. В отделительной ткани этилен вызывает образование целлюлазы. Очевидна связь этого эффекта со стимуляцией процесса отделения. Правда, ускоренное отделение наступает еще до подъема синтеза целлюлазы, но это, вероятно, объясняется тем, что этилен вызывает также высвобождение целлюлазы из связанной формы и ее секрецию в межклетники. Выделение амилазы из алейроновых клеток ячменя тоже ускоряется под действием этилена. Быстрые» эффекты этилена, например подавление клеточного растяжения, проявляющееся уже через 5 мин, связаны скорее с влиянием на мембраны, чем с изменениями белкового синтеза.[ ...]
Как известно, одной из причин токсичности почвогрунтов является их засоление. Отработанные буровые растворы и буровой шлам содержат в своем составе в ряде случаев значительное количество опасных для почв минеральных солей. Поэтому представляет интерес выявление влияния указанного фактора на биологическую продуктивность почв. Результаты исследований свидетельствуют о том, что минеральные сопи н количестве боттее 0 8-4,0 кт/м2 почвы резко снижают активность инвертазы, а в количестве более 1,5-1,6 кг/м2 почвы начинают существенно сказываться и на урожайности возделываемых на них сельхозкультур .[ ...]
Мед - высококалорийный продукт. Натуральным медом называется сладкое, вязкое и ароматическое вещество, вырабатываемое пчелами из нектара растений, а также из медвяной росы или пади. Мед может иметь вид закристаллизованной массы. Ценность меда заключается и в том, что он обладает бактерицидными свойствами. Поэтому мед не только ценный продукт питания, но и лечебное средство. Главными составными частями цветочного меда являются плодовый и виноградный сахара, которых в нем содержится около 75 %. Калорийность меда свыше 3 тыс. кал. В нем имеются ферменты: диастаза (или амилаза), инвертаза, каталаза, липаза.[ ...]
Исследования проводили в долине нижнего течения р.Сысола (Республика Коми, подзона средней тайги). Биохимические параметры почв характеризовали по уровню активности оксидоредуктаз (каталаза), гидролаз (инвертаза) и выделению С02 с поверхности почвы. Во все сроки отбора максимальные значения каталитической активности отмечены в лесных подстилках почвы Адл (4.2-8.6 мл 02/г почвы), наиболее «сухой» в исследованном ряду почв. Однако по уровню инвертазы во все сроки отбора лидировала почва Ал (11.9-37.8 мг глюкозы/г почвы в горизонте АО). В этой же почве отмечен в июле максимум в выделении С02 (0.60±0.19) кг/га-час. При использовании интегрального показателя БАП, учитывающего все параметры биологической активности, показано, что наиболее активно биологические процессы во все сроки отбора протекают в почве Ал, занимающей промежуточное положение по гидротермическому режиму между почвами Адл и Алб.[ ...]
Дестабилизация процесса нитрификации нарушает поступление в биологический круговорот нитратов, количество которых предопределяет ответную реакцию на изменение среды обитания у комплекса денитрификаторов. Ферментные системы денитрификаторов уменьшают скорость полного восстановления, слабее вовлекая закись азота в конечный этап, осуществление которого требует значительных энергетических затрат. В результате этого содержание закиси азота в надпочвенной атмосфере эродированных экосистем достигало 79 - 83% (Косинова и др., 1993). Отчуждение части органических веществ из черноземов под воздействием эрозии отражается на пополнении азотного фонда в ходе фото- и гетеротрофной фиксации азота: аэробной и анаэробной. На первых этапах эрозии быстрыми темпами идет подавление именно анаэробной азотфиксации в силу параметров лабильной части органического вещества (Хазиев, Багаутдинов, 1987). Активность ферментов инвертазы и каталазы в сильносмытых черноземах по сравнению с несмытыми уменьшилась более чем на 50%. В серых лесных почвах по мере увеличения их смытости наиболее резко снижается инвертазная активность. Если в слабосмытых почвах отмечается постепенное затухание активности с глубиной, то в сильносмытых уже в подпахотном слое инвертазная активность очень мала или не обнаруживается. Последнее связано с выходом на дневную поверхность иллювиальных горизонтов с крайне низкой активностью фермента. По активности фосфатазы и, особенно, каталазы четко выраженной зависимости от степени смытости почв не наблюдалось (Личко, 1998).[ ...]
Первичные вещества в лишайниках в общем те же, что и в других растениях. Оболочки гиф в лишайниковом слоевище составлены в основном углеводами, Часто обнаруживается в гифах хитин (С30 Н60 К4 019). Характерной составной частью гиф является полисахарид лихенин (С6Н10О6)п, называемый лишайниковым крахмалом. Реже встречающийся изомер лихенина - изолихенин - найден, кроме оболочек гиф, в протопласте. Из высокомолекулярных полисахаридов в лишайниках, в частности в оболочках гиф, встречаются гемицеллюлозы, являющиеся, очевидно, резервными углеводами. В межклеточных пространствах у некоторых лишайников обнаружены пектиновые вещества, которые, впитывая в большом количестве воду, набухают и ослизняют слоевище. В лишайниках встречаются также многие ферменты - инвертаза, амилаза, каталаза, уре-аза, зимаза, лихеназа, в том числе и внеклеточные. Из азотсодержащих веществ в гифах лишайников обнаружены многие аминокислоты - аланин, аспарагиновая кислота, глютаминовая кислота, лизин, валин, тирозин, триптофан и др. Фикобионт продуцирует в лишайниках витамины, но почти всегда в малых количествах.[ ...]
В ходе экспериментов установлено, что полужидкие и твердые отходы бурения крайне отрицательно влияют на биологическую продуктивность почв. Известно, что наибольшее негативное влияние оказывают нефть и нефтепродукты, содержащиеся в отходах . Указанные загрязнители значительно снижают активность окислительновосстановительных и гидролитических ферментов, что приводит к подавлению микробиологической активности почвы. Такой эффект ярко выражен для отходов, содержащих более 4-5 % нефти и нефтепродуктов. При меньшем содержании данного загрязнителя эффект снижения биологической продуктивности рассматриваемых типов почв характерен для периода от 3 до 6 мес., а затем отмечается усиленное размножение азотфиксирующих, денитрифицирующих и сульфатвосста-наачивающих бактерий, которые используют нефть и ее производные в качестве источника углерода и энергии, в результате чего происходят постепенное окисление и минерализация нефти. При этом закономерно падает урожайность сельхозкультур и активность инвертазы. При содержании в составе отходов более 5% нефти и нефтепродуктов видимой активности углеводородокисляющей бактериальной микрофлоры не отмечается даже по истечении 1 года. Указанный уровень загрязненности отходов является критическим, а потому требуется применение специальных агротехнических и агрохимических приемов, стимулирующих биологическую продуктивность почв (внесение удобрений, содержащих азот, фосфор и калий; интенсивная аэрация зоны нефтяного загрязнения; посев специальных трав, усиливающих деятельность углеводородусваивающей бактериальной микрофлоры) .[ ...]
Для изучения механизма и характера влияния полужидких (отработанные буровые растворы) и твердых (буровой шлам) отходов бурения, т.е. тех видов отходов, которые подвергаются засыпке минеральным грунтом в шламовых амбарах при их ликвидации, на биологическую продуктивность почв и разработки на этой основе комплекса агротехнических мер по восстановлению загрязненных земель были проведены вегетационно-полевые и полевые исследования. Эксперименты проводили по стандартным методикам . Экспериментировали с отходами бурения различной степени загрязненности по нефти и нефтепродуктам (НП), органическому углероду (показатель химического потребления кислорода - ХПК) и минеральным солям (показатель прокаленного остатка - ПО), которые добавляли в почвы в соотношении 1:1. Диапазон и уровень загрязненных отходов следующие: по НГ1 - 1,0-12,0%; по ХПК - 20,0 - 60,0 кг/м3; по ПО (в пересчете на единицу площади почвы) - 0,4-1,6 кг/м2 почвы. В исследованиях использовали три типа почвы, т.е. наиболее распространенные типы почв, на которых ведется бурение в зонах активного сельхозпользования земель. Интегральными показателями биологической продуктивности почв являлись урожайность стандартного ячменя сорта "Курьер" и активность инвертазы, которую определяли по известной методике .[ ...]
Однако, несмотря на тесную зависимость, существующую между лишайниками и субстратом, на котором они поселяются, до сих пор с достоверностью еще неизвестно, используют лишайники субстрат только как место прикрепления или они извлекают из него некоторые питательные вещества, необходимые для их жизнедеятельности. С одной стороны, способность лишайников расти на субстратах, бедных питательными веществами, дает основание считать, что они используют субстрат лишь как место прикрепления. Однако, с другой стороны, избирательная снособность, проявляемая лишайниками при расселении, строгая приуроченность большинства из них к определенному субстрату, зависимость видового состава лишайниковой растительности не только от физических, но и от химических свойств субстрата невольно наводят на мысль, что лишайники используют субстрат и как дополнительный источник питания. Это подтверждается и биохимическими исследованиями, проведенными в последние годы. Например, выяснилось, что у одного и того же вида лишайника, растущего на разных древесных породах, состав лишайниковых веществ может быть неодинаковым. Еще более очевидным доказательством служит открытие у лишайников внеклеточных ферментов, которые выделяются во внешнюю среду. Внеклеточные ферменты, такие, например, как инвертаза, амилаза, цел-люлаза и многие другие, представлены в лишайниках довольно широко и обладают достаточно высокой активностью. Причем, как оказалось, они наиболее активны в нижней части слоевища, которой лишайник прикреплен к субстрату. Это указывает на возможность активного воздействия слоевища лишайников на субстрат с целью извлечения из него дополнительных питательных веществ.
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
Введение
1. Обзор литературы
1.1 Общее представление о почвенных ферментах
1.2 Ферментативная активность почв
1.3 Методологические подходы к определению ферментативной активности почв
1.3.1 Выделение экспериментальной площади
1.3.2 Особенности отбора и подготовки образцов почв к анализу
1.4 Влияние различных факторов (температура, водный режим, сезон отбора) на ферментативную активность почв
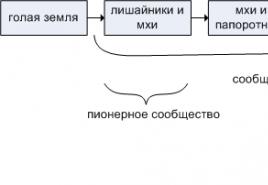
1.5 Смена сообществ микроорганизмов в почвах
1.6 Методы исследования активности почвенных ферментов
Заключение
Список использованных источников
Введение
В условиях возросшей антропогенной нагрузки на биосферу планеты, почва, являясь элементом природной системы и находясь в динамичном равновесии со всеми другими компонентами, подвергается деградационным процессам. Потоки веществ, попадая в почву в результате антропогенной деятельности, включаются в естественные циклы, нарушая нормальное функционирование почвенной биоты, и как следствие, всей почвенной системы. Среди различных биологических критериев оценки антропогенного влияния на почвы наиболее оперативными и перспективными являются биохимические показатели, дающие сведения о динамике важнейших ферментативных процессов в почве: синтезе и разложении органического вещества, нитрификации и других процессах.
Имеющиеся сведения о ферментативной активности различных типов почв в настоящее время пока недостаточны и требуют дальнейшего изучения. Это делает весьма актуальным в теоретическом и практическом отношениях изучение вопросов, затрагиваемых в данной работе .
Объект исследования: активность почвенных ферментов.
Цель курсовой работы: Изучение почвенных ферментов и ферментативной активности почв.
В соответствии с целью исследования были поставлены следующие задачи:
1. Дать общие представления о почвенных ферментах и ферментативной активности почв.
2. Рассмотреть методологические подходы к определению ферментативной активности почв.
3. Определить влияние различных природных факторов на ферментативную активность почв
4. Изучить вопрос о наличии и смене сообществ микроорганизмов в почвах
5. Перечислить и охарактеризовать методы исследования активности почвенных ферментов.
1 . Обзор литературы
1.1 Общее представление о почвенных ферментах
Трудно представить, чтобы ферменты, высокоорганизованные белковые молекулы, могли образовываться в почве вне живого организма. Известно, что почва обладает известной ферментативной активностью.
Ферменты - это катализаторы химических реакций белковой природы, отличающиеся специфичностью действия в отношении катализа определенных химических реакций.
Ферменты являются продуктами биосинтеза живых почвенных организмов: древесных и травянистых растений, мхов, лишайников, водорослей, грибов, микроорганизмов, простейших, насекомых, беспозвоночных и позвоночных животных, которые представлены в природе определенными совокупностями - биоценозами .
Биосинтез ферментов в живых организмах осуществляется благодаря генетическим факторам, ответственным за наследственную передачу типа обмена веществ и его приспособительную изменчивость. Ферменты являются тем рабочим аппаратом, при помощи которого реализуется действие генов. Они катализируют в организмах тысячи химических реакций, из которых, в итоге, слагается клеточный обмен. Благодаря ферментам химические реакции в организме осуществляются с большой скоростью.
К настоящему времени из двух тысяч известных ферментов более 150 получено в кристаллическом виде. Ферменты подразделяют на шесть классов:
1. Оксиредуктазы - катализируют окислительно-восстановительные реакции.
2. Трансферазы - катализируют реакции межмолекулярного переноса различных химических групп и остатков.
3. Гидролазы - катализируют реакции гидролитического расщепления внутримолекулярных связей.
4. Лиазы - катализирующие реакции присоединения групп по двойным связям и обратные реакции отрыва таких групп.
5. Изомеразы - катализируют реакции изомеризации.
6. Лигазы - катализируют химические реакции с образованием связей за счет АТФ (аденозинтрифосфорной кислоты).
При отмирании и перегнивании живых организмов часть их ферментов разрушается, а часть, попадая в почву, сохраняет свою активность и катализирует многие почвенные химические реакции, участвуя в процессах почвообразования и в формировании качественного признака почв - плодородия.
В разных типах почв под определенными биоценозами сформировались свои ферментативные комплексы, отличающиеся активностью биокаталитических реакций.
Важной чертой ферментативных комплексов почв является упорядоченность действия имеющихся групп ферментов. Она проявляется в том, что обеспечивается одновременное действие ряда ферментов, представляющих различные группы. Ферменты исключают накопление избытка каких-либо соединений в почве. Излишки накопившихся подвижных простых соединений (например, NH 3) тем или иным путем они временно связывают и направляют в циклы, завершающиеся образованием более сложных соединений. Ферментативные комплексы можно представить, в виде неких саморегулирующихся систем. В этом основную роль играют микроорганизмы и растения, постоянно пополняющие почвенные ферменты, так как многие из них являются короткоживущими.
О количестве ферментов косвенно судят по их активности во времени, которая зависит от химической природы реагирующих веществ (субстрата, фермента) и от условий взаимодействия (концентрация компонентов, рН, температура, состав среды, действие активаторов, ингибиторов и др.).
Ферменты, относящиеся к классам гидролаз и оксидоредуктаз, участвуют в основных процессах гумификации почв, поэтому их активность - это существенный показатель плодородия почв. Поэтому кратко остановимся на характеристике ферментов, относящихся к данным классам.
К гидролазам относят инвертазу, уреазу, фосфатазу, протеазу и др.
Инвертаза - катализирует реакции гидролитического расщепления сахарозы на эквимолярные количества глюкозы и фруктозы, воздействует также на другие углеводы (галактозы, глюкозы, рамнозы) с образованием молекул фруктозы - энергетического продукта для жизни микроорганизмов, катализирует фруктозотрансферазные реакции. Исследования многих авторов показали, что активность инвертазы лучше других ферментов отражает уровень плодородия и биологической активности почв 3, с. 27.
Уреаза - катализирует реакции гидролитического расщепления мочевины на аммиак и диоксид углерода. В связи с использованием мочевины в агрономической практике необходимо иметь в виду, что активность уреазы выше у более плодородных почв. Она повышается во всех почвах в периоды их наибольшей биологической активности - в июле-августе.
Фосфатаза (щелочная и кислая) - катализирует гидролиз ряда фосфорорганических соединений с образованием ортофосфата. Активность фосфатазы тем выше, чем меньше в почве подвижных форм фосфора, поэтому она может быть использована как дополнительный показатель при установлении потребности внесения в почвы фосфорных удобрений. Наиболее высокая фосфатазная активность в ризосфере растений.
Протеазы - это группа ферментов, расщепляющих белки до полипептидов и аминокислот, которые в последующем гидролизуются до аммиака, диоксида углерода и воды. В связи с этим протеазы имеют важнейшее значение в жизни почвы, так как с ними связаны изменение состава органических компонентов и динамика форм азота, которые легко усваиваются растениями.
К классу оксидоредуктаз относят каталазу, пероксидазу и полифенолоксидазу и др.
Каталаза - в результате ее действия происходит расщепление перекиси водорода, токсичной для живых организмов:
Н2О2 > Н2О + О2
Большое влияние на каталазную активность минеральных почв оказывает растительность. Почвы, находящиеся под растениями с мощной глубоко проникающей корневой системой, характеризуются высокой каталазной активностью. Особенность активности каталазы заключается в том, что вниз по профилю она мало изменяется, имеет обратную зависимость от влажности почв и прямую - от температуры.
Полифенолоксидазе и пероксидазе в почвах принадлежит основная роль в процессах гумусообразования.
Полифенолоксидаза катализирует окисление полифенолов в хиноны в присутствии свободного кислорода воздуха. Пероксидаза катализирует окисление полифенолов в присутствии перекиси водорода или органических перекисей. При этом ее роль состоит в активировании перекисей, поскольку они обладают слабым окисляющим действием на фенолы. Далее может происходить конденсация хинонов с аминокислотами и пептидами с образованием первичной молекулы гуминовой кислоты, которая в дальнейшем способна усложняться за счет повторных конденсаций.
Отношение активности полифенолоксидазы (S) к активности пероксидазы (D), выраженное в процентах имеет связь с накоплением в почвах гумуса, поэтому эта величина получила название условный коэффициент накопления гумуса (К):
Рассмотрим разновидности почвенных ферментов.
К классу оксидоредуктаз относятся катализирующие окислительно-восстановительные реакции.
В подавляющем большинстве биологических окислений происходит не присоединение кислорода к окисляющейся молекуле, а отщепление водорода от окисляемых субстратов. Этот процесс получил название дегидрирования и катализируется ферментами дегидрогеназами.
Различают аэробные дегидрогеназы, или оксидазы, и анаэробные дегидрогеназы, или редуктазы. Оксидазы переносят атомы водорода или электроны от окисляемого вещества на кислород воздуха. Анаэробные дегидрогеназы передают атомы водорода и электроны другим акцепторам, ферментам или переносчикам водорода, без передачи их на кислородные атомы. Окислению подвергаются многочисленные органические соединения, попадающие в почву с растениями и животными: белки, жиры, углеводы, клетчатка, органические кислоты, аминокислоты, пурины, фенолы, хиноны, специфические органические вещества типа гуминовых и фульвокислот и др.
В окислительно-восстановительных процессах в клетках животных, растений, микроорганизмов участвуют, как правило, анаэробные дегидрогеназы, которые передают отщепляемый от субстрата водород промежуточным переносчикам. В почвенной среде участвуют в окислительно-восстановительных процессах в основном аэробные дегидрогеназы, с помощью которых водород субстрата переносится непосредственно на кислород воздуха, т.е. акцептором водорода является кислород. Простейшая окислительно-восстановительная система в почвах состоит из окисляемого субстрата, оксидаз и кислорода.
Особенность оксидоредуктаз состоит в том, что, несмотря на ограниченный набор активных групп (коферментов), они способны ускорять большое число самых разнообразных окислительно-восстановительных реакций. Это достигается за счет способности одного кофермента соединяться со многими апоферментами и образовывать каждый раз оксидоредуктазу, специфичную по отношению к тому или иному субстрату.
Другой важной особенностью оксидоредуктаз является то, что они ускоряют химические реакции, связанные с высвобождением энергии, которая необходима для синтетических процессов. Окислительно-восстановительные процессы в почве катализируются как аэробными, так и анаэробными дегидрогеназами. По химической природе это двукомпонентные ферменты, состоящие из белка и активной группы, или кофермента.
В качестве активной группы могут выступать:
НАД + (никотинамидадениндинуклеотид),
НАДФ + (никотинамидадениндинуклеотидфосфат);
ФМН (флавинмононуклеотид);
ФАД (флавинадениндинуклеотид), цитохромы.
Обнаружено около пятисот различных оксидоредуктаз. Однако наиболее распространнеными оксидоредуктазами являются такие, которые содержат в качестве активной группы НАД + .
Соединяясь с белком и образуя двукомпонентный фермент (пиридинпротеин), НАД + усиливает свою способность восстанавливаться. В результате пиридинпротеины становятся способны отнимать от субстратов, в качестве которых могут быть углеводы, дикарбоновые и кетокислоты, аминокислоты, амины, спирты, альдегиды, специфические почвенные органические соединения (гуминовые и фульвокислоты) и др., атомы водорода в виде протонов (Н +). В результате активная группа фермента (НАД +) восстанавливается, а субстрат переходит в окисленное состояние.
Механизм связывания двух атомов водорода, т.е. двух протонов и двух электронов, состоит в следующем. Активный группой дегидрогеназ, которая принимает протоны и электроны, является пиридиновое кольцо. При восстановлении НАД + к одному из углеродных атомов пиридинового кольца присоединяются один протон и один электрон, т.е. один атом водорода. Второй электрон присоединяется к положительно заряженному атому азота, а оставшийся протон переходит в окружающую среду.
Все пиридинпротеины являются анаэробными дегидрогеназами. Они не передают снятые с субстрата атомы водорода на кислород, а посылают их на другой фермент.
Кроме НАД + пиридинферменты могут содержать в качестве кофермента никотинамидадениндинуклеотидфосфат (НАДФ +). Этот кофермент является производным НАД + , у которого водород ОН - группы второго углеродного атома рибозы аденозина замещен на остаток фосфорной кислоты. Механизм окисления субстратов при участии НАДФ* в качестве кофермента аналогичен НАД + .
После присоединения водорода НАДН и НАДФН обладают значительным восстановительным потенциалом. Они могут передавать свой водород другим соединениям и восстанавливать их, а сами превращаются при этом в окисленную форму. Однако водород, присоединенный к анаэробной дегидрогеназе, не может передаваться на кислород воздуха, а только преносчикам водорода. Такими промежуточными переносчиками являются флавиновые ферменты (флавопротеины). Они представляют собой двухкомпонентные ферменты, в качестве активной группы в которых может находиться фосфорилированный витамин В 2 (рибофлавин). Каждая молекула такого фермента имеет молекулу рибофлавинфосфата (или флавинмононуклеотида, ФМН). Таким образом, ФМН представляет собой соединение азотистого основания диметилизоаллоксазина с остатками пятиуглеродного спирта рибита и фосфорной кислоты. ФМН способен принимать и отдавать два атома водрода (Н) по атомам азота (N) изоаллоксазинового кольца .
Трансферазы называют ферментами переноса. Они катализируют перенос отдельных радикалов, частей молекул и целых молекул с одних соединений на другие. Реакции переноса обычно идут в две фазы. На первой фазе фермент отщепляет атомную группировку от участвующего в реакции вещества и образует с нею комплексное соединение. На второй фазе фермент катализирует присоединение группировки к другому веществу, участвующему в реакции, а сам освобождается в неизменном состоянии. Класс трансфераз насчитывает около 500 индивидуальных ферментов. В зависимости от того, какие группировки или радикалы переносят трансферазы, различают фосфотрансферазы, аминотрансферазы, гликозилтрансферазы, ацилтрансферазы, метилтрансферазы и др.
Фосфотрансферазы (киназы) - ферменты, катализирующие перенос остатков фосфорной кислоты (Н2Р03). Донором фосфатных остатков, как правило, является АТФ. Перенос фосфатных групп производится на спиртовые, карбоксильные, азотсодержащие, фосфорсодержащие и другие группы органических соединений. К фосфотрансферазам относится повсеместно распространенная гексокиназа - фермент, ускоряющий перенос остатка фосфорной кислоты от молекулы АТФ к глюкозе. С этой реакции начинается превращение глюкозы в другие соединения.
Гликозилтрансферазы ускоряют реакции переноса гликозильных остатков к молекулам моносахаридов, полисахаридов или других веществ. Это ферменты, обеспечивающие реакции синтеза новых молекул углеводов, коферментами гликозилтрансфераз являются нуклеозиддифосфатсахара (НДФ-сахара). С них в процессе синтеза олигосахаридов гликозильный остаток переносится на моносахарид. В настоящее время известно около пятидесяти НДФ-сахаров. Они широко распространены в природе, синтезируются из фосфорных эфиров моносахаридов и соответствующих нуклеозидтрифосфатов.
Ацилтрансферазы переносят остатки уксусной кислоты СН3СО - , а также остатки других жирных кислот на аминокислоты, амины, спирты и другие соединения. Это двухкомпонентные ферменты, в состав которых входит кофермент А. Источником ацильных групп является ацилкоэнзим А, который можно рассматривать как активную группу ацилтрансфераз. При переносе остатков уксусной кислоты в реакции участвует ацетил коэнзим А.
К классу гидролаз относятся ферменты, катализирующие гидролиз, а иногда и синтез сложных органических соединений с участием воды.
Подкласс эстераз включает ферменты, ускоряющие реакции гидролиза сложных эфиров, спиртов с органическими и неорганическими кислотами.
Важнейшими подподклассами эстераз являются гидролазы эфиров карбоновых кислот и фосфатазы. Реакции гидролиза жиров (триглицеридов), в результате которых освобождаются глицирин и высшие жирные кислоты, ускоряются гидролазой эфиров глицерина липазой. Различают простые липазы, которые катализируют освобождение высших жирных кислот из свободных триглицеридов, и липопротеинлипазы, осуществляющие гидролиз связанных с белками липидов. Липазы - однокомпонентные белки с молекулярным весом от 48 тыс. до 60 тыс. Хорошо изучена липаза дрожжей. Ее полипептидная цепь состоит из 430 аминокислотных остатков и сложена в глобулу, в центре которой находится активный центр фермента. Ведущую роль в активном центре липазы играют радикалы гистидина, серина, дикарбоновых кислот и изолейцина.
Активность липаз регулируется путем их фосфорилирования- дефосфорилирования. Активные липазы фосфорилированы, не активные дефосфорилированы.
Фосфатазы катализируют гидролиз фосфорных эфиров. Широко распространены фосфатазы, действующие на сложные эфиры фосфорной кислоты и углеводов. К таким соединениям относятся, например, глюкозо-6-фосфат, глюкозо-1-фосфатаза, фруктозо-1,6-дифосфат и др. Соответствующие ферменты носят названия глюкозо-6-фосфатаза, глюкозо-1-фосфатаза и т. д. Они катализируют отщепление остатка фосфорной кислоты от фосфорных эфиров:
Фосфатазы фосфодиэфиров - дезоксирибонуклеаза и рибонуклеаза катализируют расщепление ДНК и РНК до свободных нуклеотидиов.
К подклассу гидролаз относятся гликозидазы ускоряющие реакции гидролиза гликозидов. Кроме гликозидов, содержащих в качестве агликонов остатки одноатомных спиртов, субстратами, на которые действуют гликозидазы, являются олиго- и полисахариды. Из действующих на олигосахариды гликозидаз важнейшими являются мальтоза и сахароза. Они осуществляют гидролиз мальтозы и сахарозы.
Из гликозидаз, действующих на полисахариды, наибольшее значение имеют амилазы. Характерная особенность амилаз - отсутствие абсолютной специфичности действия. Все амилазы - металлопротеины, содержат Zn 2+ и Са 2+ . Активные центры амилаз образованы радикалами гистидина, аспарагиновой и глутаминовой кислот, а также тирозина. Последний выполняет функцию связывания субстрата, а первые трикаталитическую. Амилазы ускоряют реакции гидролиза гликозильных связей в молекуле крахмала с образованием глюкозы, мальтозы или олигосахаридов.
Немаловажное значение имеет целлюлаза, катализирующая расщепление целлюлозы, инулаза, расщепляющая полисахарид инулин, аглюкозидаза, превращающая дисахарид мальтозу в две молекулы глюкозы. Некоторые гликозидазы могут катализировать реакции переноса гликозильных остатков, в этом случае их называют трансгликозидазами.
Протеазы (пептидгидролазы) катализируют гидролитическое расщепление пептидных CO-NH-связей в белках или пептидах с образованием пептидов меньшей молекулярной массы или свободных аминокислот. Среди пептидгидролаз различают эндопептидазы (протеиназы), катализирующие гидролиз внутренних связей в белковой молекуле, и экзопептидазы (пептидазы), обеспечивающие отщепление от пептидной цепи свободных аминокислот.
Протеиназы делят на четыре подкласса.
1. Сериновые протеиназы, в состав активнового центра этих ферментов входит остаток серина. Последовательность аминокислотных остатков на участке полипептидной цепи у сериновых протеиназ одинакова: аспарагиновая кислота-серии-глицин. Гидроксильная группа серина характеризуется высокой процессах. Второй активной функциональной группой является имидазол остатка гистидина, активизирующий гидроксил серина в результате образования водородной связи.
2. Тиоловые (цистеиновые) протеиназы, имеют в активном центре остаток цистеина, энзиматической активностью обладают сульфгидрильные группы и ионизированная карбоксильная группа.
3. Кислые (карбоксильные) протеиназы, оптимум рН<5, содержат радикалы дикарбоновых кислот в активном центре.
4. Металлпротеиназы, каталитическое действие их обусловлено присутствием в активном центре Mg 2+, Mn 2+ , Со 2+ , Zn 2+ , Fe 2+ . Прочность связи металла с белковой частью фермента может быть различной. Входящие в состав активного центра ионы металлов принимают участие в образовании фермент-субстратных комплексов и облегчают активацию субстратов.
Важной особенностью протеиназ является селективный характер их действия на пептидные связи в белковой молекуле. В результате индивидуальный белок под воздействием определенной протеиназы расщепляется всегда на строго ограниченное число пептидов.
5. Пептидгидролазы, отщепляющие аминокислоты от пептида, начиная с аминокислоты, обладающей свободной NН2-группой, называются аминопептидазами, имеющие свободную СООН группу - карбоксипептидазами. Завершают гидролиз белка дипептидазы, расщепляя дипептиды на аминокислоты.
6. Амидазы катализируют гидролитическое расщепление связи между углеродом и азотом: дезаминирование аминов. К этой группе ферментов относится уреаза, которая осуществляет гидролитическое расщепление мочевины. окислительный фермент
7. Уреаза - однокомпонентный фермент (М=480 тыс.). Молекула представляет собой глобулу и состоит из восьми равных субъединиц. Обладает абсолютной субстратной специфичностью, действует только на мочевину .
Следует отметить, что чтобы обнаружить в почве свободные ферменты, нужно прежде всего освободить ее от живых организмов, т. е. произвести полную или частичную стерилизацию. Идеальный фактор, стерилизующий почву для нужд энзимологии, должен убивать живые клетки, не нарушая их клеточной структуры, и в то же время, не влиять на сами ферменты. Трудно сказать, все ли применяемые в настоящее время методы стерилизации отвечают этим требованиям. Чаще всего почву для нужд энзимологии стерилизуют, добавляя в качестве антисептика толуол, путем обработки почвы окисью этилена или, что теперь практикуется все чаще, убивая микроорганизмы разного рода ионизирующими излучениями. Дальнейшая техника определения каталитических свойств почвы не отличается от методов определения активности ферментов растительного или животного происхождения. К почве добавляют определенную концентрацию субстрата для фермента и после инкубации изучают продукты реакции. Анализы многих почв, проведенные этим способом, показали, что в них содержатся свободные ферменты, обладающие каталитической активностью.
1.2 Ферментативная активность почв
Ферментативная активность почв [от лат. Fermentum - закваска] -способность почвы проявлять каталитическое воздействие на процессы превращения экзогенных и собственных органических и минеральных соединений благодаря имеющимся в ней ферментам. Характеризуя ферментативную активность почв, имеют в виду суммарный показатель активности. Ферментативная активность различных почв неодинакова и связана с их генетическими особенностями и комплексом взаимодействующих экологических факторов. Уровень ферментативной активности почв определяется активностью различных ферментов (инвертазы, протеаз, уреазы, дегидрогеназ, каталазы, фосфатаз), выражаемой количеством разложенного субстрата за единицу времени на 1 г почвы.
Биокаталитическая активность почв зависит от степени обогащенности их микроорганизмами и от типа почв. Активность ферментов изменяется по генетическим горизонтам, которые отличаются по содержанию гумуса, типам реакций, окислительно-восстановительным потенциалом и другими показателями по профилю.
В целинных лесных почвах интенсивность ферментативных реакций в основном определяют горизонты лесной подстилки, а в пахотных - пахотные слои. Все биологически менее активные генетические горизонты, находящиеся под горизонтами А или Ап, имеют низкую активность ферментов. Активность их незначительно возрастает при окультуривании почв. После освоения лесных почв под пашню ферментативная активность образованного пахотного горизонта по сравнению с лесной подстилкой резко снижается, но по мере его окультуривания повышается и в сильно окультуренных почвах приближается или превышает показатели лесной подстилки.
Ферментативная активность отражает состояние плодородия почв и внутренние изменения, происходящие при сельскохозяйственном использовании и повышении уровня культуры земледелия. Эти изменения обнаруживаются как при вовлечении целинных и лесных почв в культуру, так и при различных приемах их использования .
По всей Беларуси в пахотных почвах ежегодно теряется до 0,9 т/га гумуса. В результате эрозии ежегодно безвозвратно уносится с полей 0,57 т/га гумуса. Причинами дегумификации почв являются усиление минерализации почвенного органического вещества, отставание процессов новообразования гумуса от минерализации в связи с недостаточным поступлением в почву органических удобрений и снижения ферментативной активности почвы .
Биохимические превращения органического вещества почвы происходят в результате микробиологической деятельности под влиянием ферментов. ферментативный активность почва микроорганизм
Особую роль играют ферменты в жизнедеятельности животных, растений и микроорганизмов. Почвенные ферменты участвуют при распаде растительных, животных и микробных остатков, а также синтезе гумуса. В результате питательные вещества из трудно усвояемых соединений переходят в легко доступные формы для растений и микроорганизмов. Ферменты отличаются высокой активностью, строгой специфичностью действия и большой зависимостью от различных условий внешней среды. Благодаря каталитической функции они обеспечивают быстрое протекание в организме или вне его огромного числа химических реакций .
Совместно с другими критериями ферментативная активность почв может служить надёжным диагностическим показателем для выяснения степени окультуренности почв. В результате исследований 4, с. 91 установлена зависимость между активностью микробиологических и ферментативных процессов и проведением мероприятий, повышающих плодородие почв. Обработка почв, внесение удобрений существенно изменяют экологическую обстановку развития микроорганизмов.
В настоящее время в биологических объектах обнаружено несколько тысяч индивидуальных ферментов, а несколько сотен из них выделено и изучено. Известно, что живая клетка может содержать до 1000 различных ферментов, каждый из которых ускоряет ту или иную химическую реакцию .
Интерес к применению ферментов вызван еще и тем, что постоянно возрастают требования по увеличению безопасности технологических процессов. Присутствуя во всех биологических системах, являясь одновременно продуктами и инструментами этих систем, ферменты синтезируются и функционируют при физиологических условиях (pH, температура, давление, присутствие неорганических ионов), после чего легко выводятся, подвергаясь разрушению до аминокислот. Как продукты, так и отходы большинства процессов, протекающих с участием ферментов, являются нетоксичными и легко разрушаемыми. Кроме того, во многих случаях, ферменты, используемые в промышленности, получают экологически безопасным путем. От небиологических катализаторов ферменты отличают не только безопасность и повышенная способность к биодеградации, но и специфичность действия, мягкие условия протекания реакций и высокая эффективность. Эффективность и специфичность действия ферментов позволяет получать целевые продукты с высоким выходом, что делает использование ферментов в промышленности экономически выгодным. Применение ферментов способствует сокращению расхода воды и энергии в технологических процессах, уменьшает выбросы в атмосферу CO2, снижает риск загрязнения окружающей среды побочными продуктами технологических циклов .
Применением передовой агротехники можно изменять в благоприятную сторону микробиологические процессы не только пахотного, но и подпахотного слоев почвы.
При непосредственном участии внеклеточных ферментов происходит разложение органических соединений почвы. Так, протеолитические ферменты расщепляют белковые вещества и до аминокислот.
Уреаза разлагает мочевину до СО2 и NH3. Образующийся аммиак и аммонийные соли служат источником азотного питания растений и микроорганизмов.
Инвертаза и амилаза участвуют в расщеплении углеводов. Ферменты группы фосфатов разлагают фосфорорганические соединения почвы и играют важную роль в фосфатном режиме последней.
Для характеристики общей ферментативной активности почвы обычно используют наиболее распространенные ферменты, свойственные подавляющему большинству почвенной микрофлоры - инвертазу, каталазу, протеазу и другие .
В условиях нашей республики проводилось немало исследований 16, с. 115 по изучению изменения уровня плодородия и ферментативной активности почв при антропогенном воздействии, однако полученные данные не дают исчерпывающий ответ на характер изменений из-за сложности сопоставления результатов в виду различия условий проведения опытов и методик исследований.
В связи с этим поиск оптимального решения проблемы улучшения гумусного состояния почвы и ее ферментативной активности в конкретных почвенно-климатических условиях на основе разработки ресурсосберегающих приемов основной обработки почвыё применения почвозащитных севооборотов, способствующих сохранению структуры, предотвращению переуплотнения почвы и улучшению их качественного состояния и восстановлению плодородия почв при минимальных затратах, весьма актуален.
1.3 Методологические подходы к определению ферментативной активности почв
1.3.1 Выделение экспериментально го участка и картографирование
Пробная площадка - часть исследуемой территории, характеризующаяся сходными условиями (рельефом, однородностью структуры почвы и растительного покрова, характером хозяйственного использования).
Пробная площадка должна располагаться на типичном для изучаемой территории месте. На площади 100 кв. м закладывается одна пробная площадка размером 25 м. При неоднородности рельефа площадки выбирают по элементам рельефа.
Намечают предварительный план закладки основных разрезов и полуразрезов с таким расчетом, чтобы они характеризовали почвы всех встречающихся форм рельефа местности и разностей почвенного покрова .
Способ петель применяют на территориях со сложным рельефом и густой географической сетью. При этом способе исследуемый участок расчленяют на отдельные элементарные секторы с учетом особенностей изменения рельефа или гидрографической сети. Сектор обследуют из одного центра посредством совершения петлеобразных маршрутов в радиальном направлении.
С учетом особенностей рельефа и гидрографической сети на одном конкретном участке маршруты обследования можно планировать комбинированным способом, т.е. часть участка обследуют способом параллельных пересечений территории, а часть способом петель.
По маршрутам намечают точки заложения разрезов с таким расчетом, чтобы были охвачены все основные разности рельефа и растительности, т.е. расстояния между разрезами не лимитируются, поэтому в одних, как правило, сложных по рельефу местах возможно сгущение разрезов, а на других, относительно однородных, участках расположение разрезов может быть редким.
Далее идет проведение работ, связанных с почвенным картографированием и детальным изучением почв, начинают с рекогносцировочного обследования участка (квартала). Во время рекогносцировочного обследования знакомятся с границами участка и в целом с объектом исследований, который обходят по просекам, визирам, дорогам. В наиболее характерных местах закладывают разрезы, место заложения которых наносят на план. По результатам рекогносцировочных обследований окончательно корректируют маршруты и места закладки почвенных разрезов.
После рекогносцировочного обследования приступают к собственно съемке, при выполнении которой необходимо иметь план заложения почвенных разрезов и чистую копию абриса таксационного описания. Общее представление о почвенных разностях и первоначальные засечки границ почвенных контуров получают на основе изучения основных и контрольных разрезов. Уточнение границ распространения почвенного контура производят с помощью прикопок. При этом в полевом дневнике для каждого разреза заполняют бланк описания почвенного разреза. Полевое изучение распространения почв проводят после закладки и привязки разрезов для установления классификационной принадлежности данной почвы. По результатам полевой оценки почвенного покрова и всех остальных элементов ландшафта в качестве почвенного контура выделяют обособленный, относительно однородный или однообразно-пестрый участок .
Основой для выделения границ между контурами различных почв является выявление закономерностей между почвами, рельефом и растительностью. Изменения в факторах почвообразования приводят к изменению почвенного покрова. При ясном изменении рельефа, растительных формаций и почвообразующих пород границы почвенных разностей совпадают с границами на местности. В свою очередь, легкость фиксации границ на карте и точность выделения почвенных контуров зависят от точности топографической основы. Однако в природе чаще всего приходится сталкиваться с неясными границами, постепенным переходом. В данном случае для установления границ почвенных контуров требуется заложение большого числа прикопок, а также богатый практический опыт и хорошая наблюдательность. При выполнении собственно съемки в полевых условиях, на основе скопированного из таксационного планшета плана, составляют абрис почв исследуемого участка .
Следует помнить, что строгих границ между почвенными разностями в природе не существует, так как смена одной почвенной разности другой происходит постепенно путем накопления одних признаков и утраты других. Поэтому почвенная съемка позволяет лишь в большей или меньшей мере передать схематические очертания распространения почвенных контуров, а точность выделения их границ зависит от масштаба съемки, типа почвы и других условий. Минимальные размеры почвенных контуров, подлежащих обязательному выделению на почвенной карте, определены техническими нормативами.
1.3.2 Особенности отбора и подготовки образцов почв к анализу
Чтобы правильно определить содержание того или иного вещества в почве, все агрохимические анализы должны выполняться безукоризненно точно и аккуратно. Однако даже очень тщательный анализ даст ненадежные результаты при неправильном отборе проб почвы.
Поскольку навеску для анализа берут очень маленькую, а результаты определения должны дать объективную характеристику большим количествам материала, обращают внимание на устранение неоднородности при отборе проб почвы. Усреднение почвенного образца достигается поэтапным отбором исходной, лабораторной и аналитической проб.
Смешанная исходная проба должна быть составлена из отдельных образцов (первоначальных проб), отобранных в пределах одной почвенной разности. Если участок имеет комплексный почвенный покров, то единой средней пробы взять нельзя. Их должно быть столько, сколько имеется почвенных разностей .
В зависимости от конфигурации участка расположение точек для отбора первоначальных проб на нем бывает различным. На узком, вытянутом в длину участке их можно разместить вдоль (по средине) него. На широком, близком к квадрату, участке лучше шахматное расположение мест взятия проб. На больших площадях применяется отбор почвенных проб вдоль длины участка по его средине, в количестве до 20 шт.
Взятую исходную пробу почвы тщательно перемешать на куске брезента, последовательно усредняют и уменьшают до нужного объема, затем высыпают в чистый мешочек, или коробку. Это лабораторная проба, её масса составляет около 400 г.
В коробку с лабораторной пробой сверху кладут фанерную или картонную этикетку, написанную простым карандашом, с указанием:
1. Наименования объекта.
2. Названия участка.
3. Номера делянки.
4. Глубины отбора.
5. Номера образца.
6. Фамилии руководившего работой или бравшего образец.
7. Даты проведения работы.
Такую же запись одновременно делают и в журнале.
Доставленный с участка образец почвы в лаборатории высыпают на плотную бумагу или лист чистой фанеры и разминают руками все слежавшиеся комья. Потом выбирают пинцетом посторонние включения, хорошо перемешивают почву, слегка измельчают ее. После такой подготовки лабораторной пробы ее вновь рассыпают для доведения до воздушно - сухого состояния, затем измельчают и пропускают через сито с отверстиями 2мм.
Помещение для сушки почвы должно быть сухим и защищенным от доступа аммиака, паров кислот и других газов .
Для определения ферментативной активности обычно берут почву, подсушенную на открытом воздухе; влажные образцы следует подсушивать в лаборатории при комнатной температуре. Необходимо следить за тем, чтобы образец не содержал неразложившихся растительных остатков. Комки почвы измельчают и просеивают через сито с ячеями размером 1 мм. При изучении ферментативной активности свежего (влажного) образца полному удалению растительных остатков следует уделять еще больше внимания. Одновременно с изучением активности определяют и влажность почвы, полученный результат пересчитывают на 1 г абсолютно сухой почвы.
1.4 Влияние различных факторов на ферментативную активность почв
Важным фактором, от которого зависит скорость ферментативной реакции (равно каталитическая активность фермента) является температура, влияние которой показано на рисунке 1. Из рисунка видно, что с повышением температуры до определенной величины скорость реакции увеличивается. Это можно объяснить тем, что с повышением температуры движение молекул ускоряется и у молекул реагирующих веществ оказывается больше возможности столкнуться друг с другом. Это увеличивает вероятность того, что реакция между ними произойдет. Температура, обеспечивающая наибольшую скорость реакции, называется оптимальной температурой .
Каждый фермент имеет свою оптимальную температуру. В общем для ферментов животного происхождения она лежит между 37 и 40С, а растительного - между 40 и 50С. Однако есть и исключения: б-амилаза из проросшего зерна имеет оптимальную температуру при 60С, а каталаза - в пределах 0-10С. При повышении температуры сверх оптимальной скорость ферментативной реакции снижается, хотя частота столкновений молекул увеличивается. Происходит это вследствие денатурации, т.е. потери ферментом нативного состояния. При температуре выше 80С большинство ферментов полностью теряют свою каталитическую активность.
Снижение скорости ферментативной реакции при температурах, превышающих оптимальную, зависит от денатурации фермента. Поэтому важным показателем, характеризующим отношение фермента к температуре, является его термолабильность, т.е. скорость инактивации самого фермента при повышении температуры.
Рисунок 1 - Влияние температуры на скорость гидролиза крахмала амилазой
При низких температурах (0С и ниже) каталитическая активность ферментов падает почти до нуля, но денатурация при этом не происходит. С повышением температуры их каталитическая активность вновь восстанавливается.
Также на ферментативную активность почв влияют влажность, содержание микроорганизмов, экологическое состояние почв.
1.5 Смена сообществ микроорганизмов в почвах
Микроорганизмы почвы очень многочисленны и разнообразны. Среди них имеются бактерии, актиномицеты, микроскопические грибы и водоросли, протозоа и близкие к этим группам живые существа.
Биологический круговорот в почве осуществляется с участием разных групп микроорганизмов. В зависимости от типа почвы содержание микроорганизмов колеблется. В садовых, огородных, пахотных почвах их насчитывается от одного миллиона до нескольких миллиардов микроорганизмов в 1 г почвы. В почве каждого садового участка присутствуют свои микроорганизмы. Они участвуют своей биомассой в накоплении органического вещества почвы. Они выполняют огромную роль в образовании доступных форм минерального питания растений. Исключительно велико значение микроорганизмов в накоплении биологически активных веществ в почве, таких как ауксины, гиббереллины, витамины, аминокислоты, стимулирующие рост и развитие растений. Микроорганизмы, образуют слизи полисахаридной природы, а также большое количество нитей грибов, принимают активное участие в формировании структуры почвы, склеивании пылеватых почвенных частиц в агрегаты, чем улучшают водно-воздушный режим почвы.
Биологическая активность почвы, численность и активность почвенных микроорганизмов тесно связаны с содержанием и составом органического вещества. В тоже время с деятельностью микроорганизмов тесно связаны такие важнейшие процессы формирования плодородия почв, как минерализация растительных остатков, гумификация, динамика элементов минерального питания, реакция почвенного раствора, превращения различных загрязняющих веществ в почве, степень накопления ядохимикатов в растениях, накопление токсических веществ в почве и явление почвоутомления. Велика санитарно-гигиеническая роль микроорганизмов и в трансформации и обезвреживании соединений тяжелых металлов .
Перспективным направлением восстановления и поддержания плодородия и биологической интенсификации земледелия считается применение продуктов переработки органических отходов с участием вермикомпостов дождевых червей, находящихся в симбиозе с микроорганизмами. В естественных почвах разложение опада осуществляют дождевые черви, копрофаги и другие организмы. Но в этом процессе участвуют и микроорганизмы. В кишечнике червей для них создаются более благоприятные условия для выполнения любых функций, чем в почве. Дождевые черви в союзе с микроорганизмами превращают различные органические отходы в высокоэффективные биологические удобрения с хорошей структурой, обогащенные макро- и микроэлементами, ферментами, активной микрофлорой, обеспечивающей пролонгированное (длительное, постепенное) действие на растения .
Итак, обеспечивая развитие микроорганизмов в почве, повышается урожай и улучшаете его качество. Ведь микроорганизмы развиваются, т.е. делятся каждые 20-30 мин и при наличии достаточного питания образуют большую биомассу. Если бык весом 500 кг за сутки образует 0,5 кг - 1 кг, то 500 кг микроорганизмов за сутки-биомассы, а 500 кг растений создают 5т биомассы. Почему же этого не наблюдается в почве? А потому что для этого микроорганизмам необходимо питание, а с другой стороны лимитируют различные факторы, в частности ядохимикаты. На площади 1 га в результате жизнедеятельности почвенных микробов в течение года выделяется 7500м3 углекислоты. А углекислота необходима и как источник углеродного питания растений и для растворения труднодоступных солей фосфорной кислоты и превращения фосфора в форму доступную для питания растений. Т.е. там, где хорошо работают микроорганизмы, нет необходимости во внесении фосфорных удобрений. Но сами микроорганизмы нуждаются в органическом веществе .
В балансе органического вещества почвы велика роль культурных растений. Накоплению гумуса в почвах способствует многолетние травы, особенно бобовые. После их уборки в почве остается фитомасса, которая обогащена азотом за счет фиксации его клубеньковыми бактериями из воздуха. Пропашные и овощные культуры (картофель, капуста и др.) уменьшают содержание гумуса в почве, т.к. оставляют в почве небольшое количество растительных остатков, а применяемая система глубокой обработки почвы обеспечивает интенсивное поступление в пахотный слой кислорода и, как следствие, обеспечивает сильную минерализацию органического вещества, т.е. его потерю.
При анализе почв нередко учитывается количество отдельных физиологических групп микроорганизмов. Это делается так называемым методом титра, при котором жидкие избирательные (элективные) питательные среды для определенных групп микроорганизмов заражают разными разведениями почвенной суспензии. Устанавливая после выдержки в термостате степень разведения, показавшего наличие искомой группы микроорганизмов, можно затем простым пересчетом определить численность ее представителей в почве. Таким путем узнают, насколько почва богата нитрификаторами, денитрификаторами, целлюлозоразлагающими и другими микроорганизмами.
Для характеристики типа почвы и ее состояния важны не только показатели численности разных групп микроорганизмов, но и анализ состояния в почве отдельных их видов. За редкими исключениями, даже физиологические группы микроорганизмов очень широки. Внешняя обстановка может резко менять видовой состав почвенных микроорганизмов, но мало или совсем не отражается на количестве их физиологических групп. Поэтому при анализе почвы важно стремиться установить состояние отдельных видов микроорганизмов.
Среди почвенных микроорганизмов встречаются представители разных систематических единиц, способные ассимилировать не только легкоусвояемые органические соединения, но и более сложные вещества ароматической природы, к которым относятся такие характерные для почвы соединения, как перегнойные вещества.
Все почвы на Земле образовались из выходящих на дневную поверхность весьма разнообразных горных пород, которые обычно называют материнскими. В качестве почвообразующих выступают, главным образом, рыхлые осадочные породы, так как изверженные и метаморфические породы выходят на поверхность сравнительно редко.
Основоположник научного почвоведения В. В. Докучаев рассматривал почву как особое тело природы, столь же самобытное, как растение, животное или минерал. Он указал, что в различных условиях образуются разные почвы, и что они изменяются во времени. По определению В. В. Докучаева, почвой следует называть «дневные», или поверхностные горизонты горных пород, естественно измененные влиянием ряда факторов. Тип почвы слагается в зависимости от: а) материнской породы, б) климата, в) растительности, г) рельефа страны и д) возраста почвообразовательного процесса .
Разрабатывая научные основы почвоведения, В. В. Докучаев отмечал огромную роль живых организмов, и, в частности, микроорганизмов, в формировании почвы.
Период творчества В. В. Докучаева совпал со временем великих открытий Л. Пастера, показавших огромное значение микроорганизмов в превращении разнообразных веществ и в инфекционном процессе. В конце прошлого и в начале текущего века был сделан ряд важных открытий в области микробиологии, имевших принципиальное значение для почвоведения и земледелия. Было установлено, в частности, что в почве содержится огромное количество разных микроорганизмов. Это давало повод думать о существенной роли микробиологического фактора в формировании и жизни почвы.
Одновременно с В. В. Докучаевым работал другой выдающийся ученый-почвовед П. А. Костычев 24, с. 72. В монографии «Почвы черноземной области России, их происхождение, состав и свойства» (1886) он писал, что геология имеет второстепенное значение в вопросе о черноземе, потому что накопление органического вещества происходит в верхних слоях земли, геологически разнообразных, и чернозем является вопросом географии высших растений и вопросом физиологии низших растений, разлагающих органическое вещество. П. А. Костычев провел ряд опытов по выяснению роли отдельных групп микроорганизмов в создании перегноя почвы .
Большой вклад в представления о роли биологического фактора в преобразовании Земли и в процессе почвообразования сделал ученик В. В. Докучаева академик В. И. Вернадский. Он считал, что главным фактором в миграции химических элементов в верхней части земной коры являются организмы. Их деятельность затрагивает не только органические, но и минеральные вещества почвенного и подпочвенного слоев.
Уже с начальных этапов превращения горных пород в почву роль микроорганизмов в процессах выветривания минералов вырисовывается весьма наглядно. Выдающиеся ученые В. И. Вернадский и Б. Б. Полынов рассматривали выветривание горных пород как результат деятельности растительных, преимущественно низших организмов. К настоящему времени эта, точка зрения подтверждена большим экспериментальным материалом .
Обычно первыми поселенцами горных пород являются накипные лишайники, образующие листообразные пластины, под которыми накапливается небольшое количество мелкозема. Лишайники, как правило, находятся в симбиозе с неспорообразующими сапрофитными бактериями.
В отношении ряда элементов лишайники выступают как их аккумуляторы. В мелкоземе под литофильной растительностью резко увеличивается количество органического вещества, фосфора, окиси железа, кальция и магния.
Из других растительных организмов, поселяющихся на материнских породах, следует отметить микроскопические водоросли, в частности сине-зеленые и диатомовые. Они ускоряют выветривание алюмосиликатов и также обычно живут в ассоциации с неспорообразующими бактериями.
Водоросли, очевидно, играют существенную роль как автотрофные накопители органических веществ, без которых не может протекать энергичная деятельность сапрофитных микроорганизмов. Последние продуцируют разные соединения, вызывающие выветривание минералов. Многие сине-зеленые водоросли являются фиксаторами азота и обогащают разрушаемую горную породу этим элементом.
Основную роль в процессе выветривания, вероятно, играют углекислота, минеральные и органические кислоты, вырабатываемые разными микроорганизмами. Имеются указания, что сильным растворяющим действием обладают некоторые кетокислоты. Не исключается возможность участия в выветривании перегнойных соединений.
Следует отметить, что многие бактерии образуют слизи, облегчающие тесный контакт микроорганизмов с горной породой. Разрушение последней происходит как под влиянием продуктов жизнедеятельности микроорганизмов, так и в результате образования комплексных соединений между веществом слизей и химическими элементами, входящими в состав кристаллических решеток минералов. Выветривание горных пород в природе должно рассматриваться как единство двух противоположных процессов - распада первичных минералов и возникновения вторичных минералов. Новые минералы могут возникать при взаимодействии микробные метаболитов друг с другом.
...Подобные документы
Изучение экологических условий, зональных и интразональных факторов почвообразования. Характеристика строения почвенных профилей, гранулометрического состава, физико-химических и водно-физических свойств почв, формирования агроэкологических типов почв.
курсовая работа , добавлен 14.09.2011
Характеристика морфологических элементов и признаков почвы. Типы строения почвенного профиля. Система символов для обозначения генетических горизонтов почв. Влияние химического состава на окраску почв. Классификация почвенных новообразований и включений.
реферат , добавлен 22.12.2013
Природные условия и факторы почвообразования. Систематический список основных типов почв и их морфологическая характеристика. Водно-физические свойства почв, их гранулометрический, агрегатный и химический состав, объемная масса. Методы защиты почв.
курсовая работа , добавлен 07.02.2010
Физиологическое состояние азотофиксаторов в типах почв, оценка их адаптационных возможностей. Анализ почвенных образцов, отобранных в регионах Нижегородской области. Идентификация штаммов рода Azotobacter по культуральным и физиологическим признакам.
дипломная работа , добавлен 15.02.2014
Факторы и процессы почвообразования, структура почвенного покрова объекта исследований, основные типы почв. Детальная характеристика почвенных контуров, их соотношение на исследуемой территории. Оценка плодородия почв и его лесоводческое значение.
курсовая работа , добавлен 12.11.2010
Заселение гиф Ophiobolus эубактериями, актиномицетами и грибами в естественных почвах. Антибиотическая активность некоторых особенно продуктивных грибов по отношению к другим грибам. Заражение обитающих на почве насекомых, состав бактерий в почвах.
реферат , добавлен 03.07.2011
Природные условия почвообразования: климат, рельеф, почвообразующие породы, растительность, гидрология и гидрография. Мероприятия по повышению плодородия почв, рекомендации по их использованию. Агропроизводственная группировка и бонитировка почв.
курсовая работа , добавлен 22.06.2013
Влияние пород, климата, рельефа, растительности на почвообразование. Гранулометрический состав, физические свойства, водный режим пахотных почв. Определение почвенно-экологического индекса. Основные мероприятия для повышения плодородия почв в агрогруппах.
курсовая работа , добавлен 25.05.2012
Свойства засоленных почв, их формирование. Условия аккумуляции солей в почвах. Интенсивность растительного покрова. Источники легкорастворимых солей. Распространение засоленных почв. Выражение засоленных почв в систематике, диагностические горизонты.
реферат , добавлен 30.03.2014
Изучение влияния сельскохозяйственных культур на состав и динамичность почвенных растворов. Распространение серых лесных почв, особенности генезиса, диагностика, свойства, классификация, использование. Содержание и состав органического вещества грунта.