Начальные стадии эмбриогенеза. Стадии эмбрионального развития. Мужские половые клетки - сперматозоиды
Эмбриогенез (греч. embryon - зародыш, genesis - развитие) - ранний период индивидуального развития организма от момента оплодотворения (зачатия) до рождения, является начальным этапом онтогенеза (греч. ontos - существо, genesis - развитие), процесса индивидуального развития организма от зачатия до смерти.
Развитие любого организма начинается в результате слияния двух половых клеток (гамет), мужской и женской. Все клетки тела, несмотря на различия в строении и выполняемых функциях, объединяет одно - единая генетическая информация, хранящаяся в ядре каждой клетки, единый двойной набор хромосом (кроме узкоспециализированных клеток крови - эритроцитов, которые не имеют ядра). То есть, все соматические (сома - тело) клетки диплоидны и содержат двойной набор хромосом - 2 n, и лишь половые клетки (гаметы), формирующиеся в специализированных половых железах (семенниках и яичниках), содержат одинарный набор хромосом - 1 n.
При слиянии половых клеток образуется клетка - зигота, в которой восстанавливается двойной набор хромосом. Напомним, что в ядре клетки человека содержится 46 хромосом, соответственно половые клетки имеют 23 хромосомы
Образовавшаяся зигота начинает делиться. I этап деления зиготы называется дроблением, в результате которого образуется многоклеточная структура морула (тутовая ягода). Цитоплазма распределяется между клетками неравномерно, клетки нижней половины морулы крупнее, чем верхней. По объему морула сравнима с объемом зиготы.
На II этапе деления, в результате перераспределения клеток, образуется однослойный зародыш - бластула, состоящий из одного слоя клеток и полости (бластоцель). Клетки бластулы различаются между собой по размерам.
На III этапе, клетки нижнего полюса как бы впячиваются (инвагинируют) вовнутрь, и образуется двухслойный зародыш - гаструла, состоящий из наружного слоя клеток - эктодермы и внутреннего слоя клеток - энтодермы.
Очень скоро, между I и II слоями клеток формируется, в результате деления клеток, еще один слой клеток, средний - мезодерма, и зародыш становится трехслойным. На этом завершается стадия гаструлы.
Из этих трех слоев клеток (их называют зародышевыми слоями) формируются ткани и органы будущего организма. Из эктодермы развивается покровная и нервная ткань, из мезодермы - скелет, мышцы, кровеносная система, половые органы, органы выделения, из энтодермы - органы дыхания, питания, печень, поджелудочная железа. Многие органы формируются из нескольких зародышевых слоев.
Эмбриогенез включает в себя процессы с момента оплодотворения до рождения..
Развитие человеческого организма начинается после оплодотворения женской половой клетки – яйца (ovium) мужской – сперматозоидом (spermatozoon, spermium).
Детальное изучение развития человеческого зародыша (эмбриона) составляет предмет эмбриологии. Здесь мы ограничимся лишь общим обзором развития зародыша (эмбриогенеза), что необходимо для понимания телосложения человека.
Эмбриогенез всех позвоночных, в том числе и человека, можно разделить на три периода.
1. Дробление: оплодотворенное яйцо, spermovium, или зигота последовательно делится на клетки (2,4,8,16 и так далее) в результате чего сначала образуется плотный многоклеточный шар, морула, а затем однослойный пузырек – бластула, которая содержит в середине первичную полость, бластоцель. Длительность этого периода – 7 дней.
2. Гаструляция заключается в превращении однослойного зародыша в двох-, а позже трехслойный – гаструлу. Первые два слоя клеток называются зародышевыми листками: внешний эктодерма и внутренний энтодерма (до двух недель после оплодотворения), а возникающий позже между ними третий, средний, слой получает название среднего зародышевого листка - мезодермы. Вторым важным результатом гаструляции у всех хордовых является возникновение осевого комплекса зачатков: на дорсальной (спинной) стороне энтодермы возникает зачаток спинной струны, хорды, а на вентральной (брюшной) ее стороне – зачаток кишечной энтодермы; на дорзальной стороне зародыша, по средней линии его из эктодермы выделяется нервная пластинка – зачаток нервной ситеми, а остальная эктодерма идет на построение эпидермиса кожи и потому называется кожной эктодермой.
В дальнейшем зародыш растет в длину и превращается в цилиндрическое образование с головным (краниальним) и хвостовым каудальным концами. Этот период длится до конца третьей недели после оплодотворения.
3. Органогенез и гистогенез: нервная пластинка погружается под эктодерму и превращается в нервную трубку, которая состоит из отдельных сегментов – невротомов, – и дает начало развитию нервной системы. Мезодермальные зачатки отшнуровываются от энтодермы первичной кишки и образуют парной ряд метамерно размещенных мешков, которые, разрастаясь по бокам от тела зародыша, делятся каждый на два отдела: спинной, что лежит по бокам от хорды и нервной трубки, и брюшной, что лежит по бокам от кишки. Спинные отделы мезодермы образуют первичные сегменты тела – сомиты, каждый из которых в свою очередь делится на склеротом, который дает начало скелету и миотом, из которого развивается мускулатура. Из сомита (на боковой его стороне) выделяется также кожный сегмент – дерматом. Брюшные отделы мезодермы, которые называются спланхнотомами, образуют парные мешки, которые содержат вторичную полость тела.
Кишечная энтодерма, которая осталась после обособления хорды и мезодермы, образует вторичную кишку – основание для развития внутренних органов. В последующем закладываются все органы тела, материалом для построения которых служат три зародышевых листка.
1. Из внешнего зародышевого листка, эктодермы, развиваются:
а) эпидермис кожи и его производные (волосы, ногти, кожные железы);
б) эпителий слизистой оболочки носа, рта и заднего прохода;
в) нервная система и эпителий органов чувств.
2. Из внутреннего зародышевого листка, энтодермы, развивается эпителий слизистой большей части пищеварительного тракта со всеми принадлежащими сюда железистыми структурами, большей части дыхательных органов, а также эпителий щитовидной и зобной желез.
3. Из среднего зародышевого листка, мезодермы, развивается мускулатура скелета, мезотелий облочек серозных полостей с зачатками половых желез и почек.
Кроме того, из спинных сегментов мезодермы возникает эмбриональная соединительная ткань, мезенхима, которая дает все виды соединительной ткани, в том числе хрящевую и костную. Так как сначала мезенхима проводит питательные вещества к разным участкам зародыша, выполняя трофическую функцию, то позже из нее развиваются кровь, лимфа, кровеносные сосуды, лимфатические узлы, селезенка.
Кроме развития самого зародыша, необходимо учитывать также образование внезародышевых частей, с помощью которых эмбрион получает необходимые для его жизни питательные вещества.
В многоклеточном плотном шаре выделяется внутренний зародышевый узелок, ембриобласт, и внешний слой клеток, который играет важную роль в питании зародыша и потому называется трофобластом. С помощью трофобласта зародыш проникает в толщу слизистой оболочки матки (вживление), и здесь начинается образование особенного органа, с помощью которого устанавливается связь зародыша с телом матери и осуществляется его питание. Этот орган называется детским местом, пометом, или плацентой. Млекопитающие, которые имеют плаценту называются плацентарными. Рядом с образованием плаценты идет процесс обособления зародыша, который развивается, от внезародышевых частей в результате возникновения так называемой туловищной складки, которая, вдаваясь гребнем к середине, будто отшнуровывает кольцом тело зародыша от внезародышевых частей. При этом, однако, сохраняется соединение с плацентой с помощью пупочного стебля, который дальше превращается в пупочный канатик. На ранних стадиях развития в последнем проходит желточная протока, которая соединяет кишку с ее выпячиванием в внезародышевый участок, – желточный мешок. У позвоночных, которые не имеют плаценты желточный мешок содержит питательный материал яйца – желток и является важным органом, через который осуществляется питание зародыша.
У человека желточный мешок хотя и возникает, но заметную роль в развитии зародыша не играет и после всасывания его содержимого постепенно редуцируется. В пупочном канатике проходят также пупочные (плацентарные) сосуды, через которые течет кровь от плаценты в тело зародыша и назад. Они развиваются из мезодермы мочевого мешка, или алантоиса, который выпирается из вентральной стенки кишки и выходит из тела зародыша через пупочное отверстие во внезародышевую часть. У человека из части алантоиса, что содержится в середине тела зародыша, образуется часть мочевого пузыря, а из его сосудов образуются пупочные кровеносные сосуды. Зародыш, который развивается, покрыт двумя зародышевыми оболочками. Внутренняя оболочка, амнион, образует объемитстий мешок, который наполнен белковой жидкостью и образует жидкую среду для зародыша, через что мешок называют водной оболочкой. Весь зародыш вместе с амниотичным и желточным мешками окружен внешней оболочкой (в состав которой входит и трофобласт). Эта оболочка, имея ворсинки, называется ворсинчатой, или хорион. Хорион выполняет трофическую, дыхательную, выделительную и барьерную функции.
13. В отличие от мхов, у папоротников, хвощей и плаунов в цикле разви тия преобладает спорофит - листостебельное растение. Представители этих трех групп растений имеют листья, стебли и корни. У большинства из них имеются подземные корневища с видоизмененными листьями и придаточными корнями. Современные хвощи, плауны и папоротники - в основном травянистые растения. Только в тропиках и субтропиках рас тут древовидные папоротники. Однако в древние эпохи - 200-350 млн лет назад эти группы растений были представлены древовидными форма ми и составляли дремучие леса, давшие начало крупнейшим каменно
угольным отложениям мира (Донбасс, Кузбасс и др.).
Каковы особенности строения плаунов, хвощей и папоротников?
Рассмотрим особенности плаунов, хвощей и папоротников. Современные плауновидные - многолетние, обычно вечнозеленые тра вы. Самый известный представитель плауновидных - плаун обыкновен ный, распространенный в средней полосе России в сыроватых еловых и
сосновых лесах. Это растение с гибким разветвленным стеблем, стелющим ся по земле. Листья мелкие, расположены на стебле по спирали. В конце лета на боковых веточках появляются обычно два спороносных колоска. Каждый колосок образован мелкими тонкими видоизмененными листья ми, которые называются спорофиллами. В основании спорофиллов распо ложены спорангии, где формируются споры. Хвощи, или хвощевидные, легко отличить по членистому строению стеблей: у них ярко выражено чередование узлов и междоузлий. Листья на стебле располагаются мутовками (по несколько штук в узле), окружая стебель. На верхушках стеблей формируются спороносные колоски, в ко торых созревают споры. У некоторых видов, например у хвоща полевого, стебли бывают двух типов: спороносные (буровато розовые, развиваются весной и после спороношения отмирают) и вегетативные (появляются летом от того же самого корневища). Папоротниковидные представлены в природе многолетними травами, лианами, деревьями и эпифитами, поселяющимися на стволах деревьев. У папоротников крупные листья; молодые обычно свернуты в виде улит ки. Папоротники нашей страны имеют корневища. Спорангии у них располагаются на нижней стороне листа и собраны в кучки - их называ ют сорусами. У вымерших папоротников спорангии были одиночными. На территории нашей страны растут папоротники орляк обыкновенный, щи товник мужской, кочедыжник женский, многоножка обыкновенная и дру гие виды.
В чем особенность развития папоротников, хвощей и плаунов?
Размножение у всех трех групп высших споровых растений происхо дит по одной схеме. Рассмотрим его на примере папоротника. На нижней стороне листа взрослого растения развиваются спорангии со спо рами. Попадая в благоприятные условия, спора прорастает и дает начало гаметофиту. Он имеет вид маленькой пластинки с ризоидами и называет ся заростком. На заростке развиваются мужские и женские гаметангии с
половыми клетками - яйцеклетками и сперматозоидами. После оплодо творения, которое происходит при наличии воды, из зиготы сначала раз вивается зародыш, а потом и взрослое растение - спорофит. Таким образом, у плаунов, хвощей и папоротников имеет место чере дование полового поколения (заросток - гаметофит) и бесполого (взрос
лое растение - спорофит).
14. Послезародышевое развитие: прямое и непрямое. Причины ослабления конкуренции между родителями и потомством при непрямом развитии.
1. Индивидуальное развитие организма (онтогенез) - период жизни, который при половом размножении начинается с образования зиготы, характеризуется необратимыми изменениями (увеличением массы, размеров, появлением новых тканей и органов) и завершается смертью.
2. Зародышевый (эмбриональный) и послезаро-дышевый (постэмбриональный) периоды индивидуального развития организма.
3. Послезародышевое развитие (приходит на смену зародышевому) - период от рождения или выхода зародыша из яйца до смерти. Различные пути послезародышевого развития животных - прямое и непрямое:
1) прямое развитие - рождение потомства, внешне похожего на взрослый организм. Примеры: развитие рыб, пресмыкающихся, птиц, млекопитающих, некоторых видов насекомых. Так, малек рыбы похож на взрослую рыбу, утенок на утку, котенок на кошку;
2) непрямое развитие - рождение или выход из яйца потомства, отличающегося от взрослого организма по морфологическим признакам, образу жизни (типу питания, характеру передвижения). Пример: из яиц майского жука появляются червеобразные личинки, живут в почве и питаются корнями в отличие от взрослого жука (живет на дереве, питается листьями).
Стадии непрямого развития насекомых: яйцо, личинка, куколка, взрослая особь. Особенности жизни животных на стадии яйца и куколки - они неподвижны. Активный образ жизни личинки и взрослого организма, разные условия обитания, использование разной пищи.
4. Значение непрямого развития - ослабление конкуренции между родителями и потомством, так как они поедают разную пищу, у них разные места обитания. Непрямое развитие - важное приспособление, возникшее в процессе эволюции. Оно способствует ослаблению борьбы за существование между родителями и потомством, выживанию животных на ранних стадиях послезародышевого развития.
Общая характеристика. Первые голосеменные появились в конце девонского периода около 350 млн. лет назад; вероятно, они произошли от древних папоротниковидных, вымерших в начале каменноугольного периода. В мезозойскую эру - эпоху горообразования, поднятия материков и иссушения климата - голосеменные достигли расцвета, но уже с середины мелового периода уступили свое господствующее положение покрытосеменным.
Отдел современных голосеменных насчитывает более 700 видов. Несмотря на относительно малую численность видов, голосеменные завоевали почти весь земной шар. В умеренных широтах Северного полушария они на огромных пространствах образуют хвойные леса, называемые тайгой.
Современные голосеменные представлены преимущественно деревьями, значительно реже - кустарниками и очень редко -лианами; травянистых растений среди них нет. Листья голосеменных значительно отличаются от других групп растений не только по форме и размерам, но и по морфологии и анатомии. У большинства видов они игловидные (хвоя) или чешуевидные; у отдельных представителей они крупные (например, у вельвичии удивительной их длина достигает 2-3 м), перисторассеченные, двулопастные и др. Листья располагаются поодиночке, по два или несколько в пучках.
Подавляющее большинство голосеменных - вечнозеленые, одно- или двудомные растения с хорошо развитыми стеблем и корневой системой, образованной главным и боковым корнями. Расселяются они семенами, которые формируются из семязачатков. Семязачатки голые (отсюда название отдела), расположены на мегаспорофиллах или на семенных чешуях, собранных в женские шишки.
В цикле развития голосеменных наблюдается последовательная смена двух поколений - спорофита и гаметофита с господством спорофита. Гаметофиты сильно редуцированы, причем мужские гаметофиты голо- и покрытосеменных растений не имеют антеридиев, чем резко отличаются от всех разноспоровых бессеменных растений.
Голосеменные включают шесть классов, два из которых полностью исчезли, а остальные представлены ныне живущими растениями. Наиболее сохранившейся и самой многочисленной группой голосеменных является класс Хвойные, насчитывающий не менее 560 видов, образующих леса на обширных пространствах Северной Евразии и Северной Америки. Наибольшее число видов сосны, ели, лиственницы встречается у побережий Тихого океана.
Класс Хвойные. Все хвойные - вечнозеленые, реже листопадные (например, лиственница) деревья или кустарники с игольчатыми или чешуевидными {например, у кипариса) листьями. Игловидные листья (хвоя) плотные, кожистые и жесткие, покрыты толстым слоем кутикулы. Устьица погружены в углубления, заполненные воском. Все эти особенности строения листьев обеспечивают хорошее приспособление хвойных к произрастанию как в засушливых, так и в холодных местообитаниях.
У хвойных прямостоячие стволы, покрытые чешуйчатой корой. На поперечном разрезе стебля хорошо видны развитая древесина и менее развитые кора и сердцевина. Ксилема хвойных на 90-95% образована трахеидами. Шишки хвойных раздельнополые; растения - чаще однодомные, реже - двудомные.
Наиболее широко распространенными представителями хвойных в Беларуси и России являются сосна обыкновенная и ель обыкновенная, или европейская. Их строение, размножение, чередование поколений в цикле развития отражает характерные особенности всех хвойных.
Сосна обыкновенная -однодомное растение (рис. 9.3). В мае у основания молодых побегов сосны образуются пучки зеленовато-желтых мужских шишек длиной 4-6 мм и диаметром 3-4 мм. На оси такой шишки расположены многослойные чешуйчатые листочки, или микроспорофиллы. На нижней поверхности микроспорофиллов находятся два микроспорангия - пыльцевых мешка, в которых образуется пыльца. Каждое пыльцевое зерно снабжено двумя воздушными мешками, что облегчает перенос пыльцы ветром. В пыльцевом зерне имеются две клетки, одна из которых впоследствии, при попадании на семязачаток, формирует пыльцевую трубку, другая после деления образует два спермия.
На других побегах того же растения образуются женские шишки красноватого цвета. На их главной оси располагаются мелкие прозрачные кроющие чешуйки, в пазухах которых сидят крупные толстые, впоследствии одревесневающие чешуи. На верхней стороне этих чешуй расположено по два семязачатка, в каждом из которых развивается женский гаметофит - эндосперм с двумя архегониями с крупной яйцеклеткой в каждом из них. На верхушке семязачатка, снаружи защищенного интегументом, имеется отверстие - пыльцевход, или микропиле.
Поздней весной или в начале лета созревшая пыльца разносится ветром и попадает на семязачаток. Через микропиле пыльца втягивается внутрь семязачатка, где и прорастает в пыльцевую трубку, которая проникает к архегониям. Образовавшиеся к этому времени два спермия по пыльцевой трубке попадают к архегониям. Затем один из спермиев сливается с яйцеклеткой, а другой отмирает. Из оплодотворенной яйцеклетки (зиготы) формируется зародыш семени, а семязачаток превращается в семя. Семена у сосны созревают на второй год, высыпаются из шишек и, подхваченные животными или ветром, переносятся на значительные расстояния.
По своему значению в биосфере и роли в хозяйственной деятельности человека хвойные занимают второе место после покрытосеменных, далеко превосходя все остальные группы высших растений.
Они помогают решать огромные водоохранные и ландшафтные задачи, служат важнейшим источником древесины, сырья для получения канифоли, скипидара, спирта, бальзамов, эфирных масел для парфюмерной промышленности, лекарственных и других ценных веществ. Некоторые хвойные культивируются как декоративные (пихты, туи, кипарисы, кедры и др.). Семена ряда сосен (сибирской, корейской, итальянской) употребляются в пищу, из них также получают масло.
Представители других классов голосеменных (саговниковые, гнетовые, гинкговые) встречаются значительно реже и менее известны, чем хвойные. Однако почти все виды саговниковых декоративны и пользуются широкой популярностью у садовников многих стран. Вечнозеленые безлистные невысокие кустарники эфедры (класс гнетовых) служат источником сырья для получения алкалоида эфедрина, который применяется как средство, возбуждающее центральную нервную систему, а также при лечении заболеваний аллергического характера.
16. Радиальная симметрия - форма симметрии, при которой тело (или фигура) совпадает само с собой при вращении объекта вокруг определённой точки или прямой. Часто эта точка совпадает с центром симметрии объекта, то есть той точкой, в которой пересекается бесконечное количество осей двусторонней симметрии. Радиальной симметрией обладают такие геометрические объекты, как круг, шар, цилиндр или конус.
В биологии о радиальной симметрии говорят, когда через трёхмерное существо проходят одна или более осей симметрии. При этом радиальносимметричные животные могут и не иметь плоскостей симметрии. Так, у сифонофоры Velella имеется ось симметрии второго порядка и нет плоскостей симметрии
Обычно через ось симметрии проходят две или более плоскости симметрии. Эти плоскости пересекаются по прямой - оси симметрии. Если животное будет вращаться вокруг этой оси на определённый градус, то оно будет отображаться само на себе (совпадать само с собой).
Таких осей симметрии может быть несколько (полиаксонная симметрия) или одна (монаксонная симметрия). Полиаксонная симметрия распространена среди протистов (например, радиолярий).
Как правило, у многоклеточных животных два конца (полюса) единственной оси симметрии неравноценны (например, у медуз на одном полюсе (оральном) находится рот, а на противоположном (аборальном) - верхушка колокола. Такая симметрия (вариант радиальной симметрии) в сравнительной анатомии называется одноосно-гетеропольной. В двухмерной проекции радиальная симметрия может сохраняться, если ось симметрии направлена перпендикулярно к проекционной плоскости. Иными словами, сохранение радиальной симметрии зависит от угла наблюдения.
Радиальная симметрия характерна для многих стрекающих, а также для большинства иглокожих. Среди них встречается так называемая пентасимметрия, базирующаяся на пяти плоскостях симметрии. У иглокожих радиальная симметрия вторична: их личинки двустороннесимметричны, а у взрослых животных наружная радиальная симметрия нарушается наличием мадрепоровой пластинки.
Кроме типичной радиальной симметрии существует двулучевая радиальная симметрия (две плоскости симметрии, к примеру, у гребневиков). Если плоскость симметрии только одна, то симметрия билатеральная (такую симметрию имеют животные из группы Bilateria).
У цветковых растений часто встречаются радиальносимметричные цветки: 3 плоскости симметрии (водокрас лягушачий), 4 плоскости симметрии (лапчатка прямая), 5 плоскостей симметрии (колокольчик), 6 плоскостей симметрии (безвременник). Цветки с радиальной симметрией называются актиноморфные, цветки с билатеральной симметрией - зигоморфные.
Билатера́льная симме́трия (двусторонняя симметрия) - симметрия зеркального отражения, при которой объект имеет одну плоскость симметрии, относительно которой две его половины зеркально симметричны. Если на плоскость симметрии опустить перпендикуляр из точки A и затем из точки О на плоскости симметрии продолжить его на длину AО, то он попадёт в точку A1, во всём подобную точке A. Ось симметрии у билатерально симметричных объектов отсутствует. У животных билатеральная симметрия проявляется в схожести или почти полной идентичности левой и правой половин тела. При этом всегда существуют случайные отклонения от симметрии (например, различия в папиллярных линиях, ветвлении сосудов и расположении родинок на правой и левой руках человека). Часто существуют небольшие, но закономерные различия во внешнем строении (например, более развитая мускулатура правой руки у праворуких людей) и более существенные различия между правой и левой половиной тела в расположении внутренних органов. Например, сердце у млекопитающих обычно размещено несимметрично, со смещением влево.
У животных появление билатеральной симметрии в эволюции связано с ползанием по субстрату (по дну водоема), в связи с чем появляются спинная и брюшная, а также правая и левая половины тела. В целом среди животных билатеральная симметрия более выражена у активно подвижных форм, чем у сидячих. Билатеральная симметрия свойственна всем достаточно высокоорганизованным животным, кроме иглокожих. В других царствах живых организмов билатеральная симметрия свойственна меньшему числу форм. Среди протистов она характерна для дипломонад (например, лямблий), некоторых форм трипаносом, бодонид, раковинок многих фораминифер. У растений билатеральную симметрию имеет обычно не весь организм, а его отдельные части - листья или цветки. Билатерально симметричные цветки ботаники называют зигоморфными.
17. Покрытосеменные (цветковые, пестичные) по времени появления на Земле являются самой молодой и в то же время наиболее высокоорганизованной группой растений. В процессе эволюции представители этого отдела появились позднее других, но они очень быстро заняли господствующее положение на земном шаре.
Наиболее характерной отличительной особенностью покрытосеменных является наличие у них своеобразного органа - цветка, который отсутствует у представителей других отделов растений. Поэтому покрытосеменные и называются чаще еще цветковыми растениями. Семяпочка у них скрытая, она развивается внутри пестика, в его завязи, поэтому покрытосеменные называются иначе пестичными. Пыльца у покрытосеменных улавливается не семяпочками, как у голосеменных, а особым образованием - рыльцем, которым заканчивается пестик.
После оплодотворения яйцеклетки из семяпочки образуется семя, а завязь разрастается в плод. Следовательно, семена у покрытосеменных развиваются в плодах, поэтому этот отдел растений и называется покрытосеменные.
Покрытосеменные (Angiospermae), или цветковые (Magnoliophyta) – отдел наиболее совершенных высших растений, имеющих цветок. Ранее включались в отдел семенных растений вместе с голосеменными. В отличие от последних семязачатки цветковых заключены в завязь, образованную сросшимися плодолистиками.
Цветок является генеративным органом покрытосеменных растений. Он состоит из цветоножки и цветоложа. На последнем располагаются околоцветник (простой или двойной), андроцей (совокупность тычинок) и гинецей (совокупность плодолистиков). Каждая тычинка состоит из тонкой тычиночной нити и расширенного пыльника, в котором созревают спермии. Плодолистик цветковых растений представлен пестиком, который состоит из массивной завязи и длинного столбика, вершинная расширенная часть которого называется рыльце.
Покрытосеменные имеют вегетативные органы, обеспечивающие механическую опору, транспорт, фотосинтез, газообмен, а также запасание питательных веществ, и генеративные органы, участвующие в половом размножении. Внутреннее строение тканей наиболее сложно из всех растений; ситовидные элементы флоэмы окружены клетками-спутницами; почти все представители покрытосеменных имеют сосуды ксилемы.
Содержащиеся внутри пыльцевых зёрен мужские гаметы попадают на рыльце и прорастают. Гаметофиты цветковых крайне упрощены и миниатюрны, что значительно сокращает длительность цикла размножения. Образуются они в результате минимального количества митозов (трёх у женского гаметофита и двух у мужского). Одна из особенностей полового размножения – двойное оплодотворение, когда один из спермиев сливается с яйцеклеткой, образуя зиготу, а второй – с полярными ядрами, образуя эндосперм, служащий запасом питательных веществ. Семена цветковых растений заключены в плод (отсюда их второе название – покрытосеменные).
18. ДНК Явление это было открыто в опытах с пневмококками, то есть с бактериями, вызывающими воспаление легких. Известны две формы пневмококков: А-форма с полисахаридной капсулой и Б-форма без капсулы. Оба эти признака наследственны.
Пневмококки А-формы при заражении ими мышей вызывают воспаление легких, от которого мыши погибают. Б-форма для них безвредна.
В 1928 году английский бактериолог Ф. Гриффитс заражал мышей смесью, состоящей из убитых нагреванием пневмококков А-формы и живых пневмококков Б-формы. Ученый предполагал, что мыши не заболеют. Но вопреки ожиданиям подопытные животные погибли. Ф. Гриффитсу удалось выделить из тканей погибших мышей пневмококки. Все они оказались капсулированными, то есть А-формы. Следовательно, убитая форма каким-то образом передавала свои свойства живым клеткам Б-формы. Но как? С помощью какого именно вещества: полисахарида, из которого состоит капсула, белка или ДНК?
От решения этого вопроса зависело многое, так как, установив вещество, передающее наследственный признак - образование капсулы, можно было получить нужный ответ. Однако сделать это не удавалось довольно долго. Лишь спустя 16 лет после опытов Ф. Гриффитса, в 1944 году, американский ученый А. Эвери с сотрудниками, поставив ряд четких экспериментов, сумел с полным обоснованием доказать, что полисахарид и белок не имеют никакого отношения к передаче наследственных свойств пневмококка А-формы.
В процессе этих экспериментов с помощью специального фермента растворили полисахаридную капсулу убитых пневмококков А-формы и проверили, продолжают ли остатки клетки формы А передавать наследственную информацию клеткам формы Б. Оказалось, что продолжают. Стало ясно, что полисахарид как источник генетической информации отпадает.
Таким образом, методом исключения было установлено, что наследственную информацию в клетке хранит и передает молекула ДНК. И действительно, когда разрушили ДНК, образование капсульных форм А из бескапсульных Б прекратилась.
Явление преобразования, то есть наследственного изменения свойств одной формы бактерий под воздействием веществ другой формы, было названо трансформацией. Вещество же, вызывающее трансформацию, получило название трансформирующего агента. Им, как было установлено, служит ДНК.
Каждый белок представлен одной или несколькими полипептидными цепями. Участок ДНК, несущий информацию об одной полипептидной цепи, называется геном. Каждая молекула ДНК содержит множество разных генов. Совокупность молекул ДНК клетки выполняет функцию носителя генетической информации. Благодаря уникальному свойству - способности к удвоению, которым не обладает ни одна другая из известных молекул, ДНК могут копироваться. При делении "копии" ДНК расходятся по двум дочерним клеткам, каждая из которых вследствие этого будет иметь ту же информацию, которая содержалась в материнской клетке. Так как гены - это участки молекул ДНК, то две клетки, образующиеся при делении, имеют одинаковые наборы генов. Каждая клетка многоклеточного организма при половом размножении возникает из одной оплодотворенной яйцеклетки в результате многократных делений. Значит, случайно возникшая ошибка в гене одной клетки будет воспроизведена в генах миллионов ее потомков. Вот почему все эритроциты больного серповидноклеточной анемией имеют одинаково испорченный гемоглобин. Ошибка произошла в гене, несущем информацию о бета-цепи белка. Копией гена является и-РНК. По ней, как по матрице, в каждом эритроците тысячи раз "печатается" неправильный белок. Дети получают испорченные гены от родителей через их половые клетки. Генетическая информация передается как от одной клетки в дочерние клетки, так и от родителей детям. Ген является единицей генетической, или наследственной, информации.
Эмбриология
(от греч. Embryon - зародыш и logos - слово, наука) - наука о развитии зародыша. Она изучает период индивидуального развития, который начинается оплодотворением и заканчивается (у млекопитающих и человека) рождением плода.
Эмбриология изучает морфологические проявления образования из оплодотворенной яйцеклетки (зиготы) сложного многоклеточного организма, в состав которого входят четыре ткани, несколько десятков клеточных диферонов, органы, системы органов и аппараты и части тела.
Знания по эмбриологии способствуют лучшему усвоению макроскопической анатомии. Выстраиваясь в логический ряд с эмбриологических данным, анатомические факты, касающиеся макро-и микроструктуры организма, подтверждают, что структурные зависимости в постнатальном периоде развития - закономерный результат процессов морфогенеза, происходящих в зародышах.
Эмбриология позволяет определить морфологические проявления, полноценность реализации генетической программы, заложенной в зиготе, а также нарушение этой реализации, что проявляется в структурных и функциональных аномалиях эмбрионального развития, которые нередко встречаются в клинической практике и объяснить которые можно, только зная эмбриологию.
Процесс эмбрионального развития человека, как и других позвоночных, делится на ряд стадий, характеризующихся качественными и количественными особенностями. Главными стадиями эмбриогенеза являются: оплодотворение, дробление, гаструляция, образование тканей (гистогенез), органов (органогенез), а также систем органов и аппаратов (системогенеза). В некоторых учебниках и пособиях до эмбриогенеза относят также прогенез - развитие женских и мужских половых клеток.
В этом разделе рассмотрены, главным образом, начальные этапы развития зародыша человека, который начинается оплодотворением и завершается образованием зачатков тканей и внезародышевых органов. Что касается гисто-, органо-и системогенеза, то они будут рассматриваться при изучении строения соответствующих органов и систем.
Оплодотворение представляет собой слияние мужской и женской половых клеток, что приводит к образованию одноклеточного зародыша - зиготы, содержащей диплоидный набор хромосом и объединяет материнскую и отцовскую наследственность. Оплодотворение происходит в яйцеводах. Продолжительность процесса - около суток.
Зигота начинает делиться путем митоза. Ее разделение называется дроблением, ибо в процессе последовательного деления зиготы клетки уменьшаются. В связи с этим общие размеры многоклеточного зародыша поначалу могут быть примерно такими же, как и зиготы.
Клетки, образующиеся в процессе дробления, получили название бластомеров (от греч. Blastos - зачаток, meros - часть). Период дробления начинается с деления зиготы на два бластомера и продолжается вплоть до образования пузырька (бластулы). У человека этот период начинается через сутки после оплодотворения и длится примерно 4-5 суток.
Одна из особенностей дробления зиготы млекопитающих и человека - возникновение бластомеров, которые различаются между собой величиной и скоростью дробления. Внешне расположены мелкие, светлые бластомеры (микромеры), что скорее делятся и образуют в совокупности трофобласт, или Живного зачаток (от греч. Tropho - питаю). В центре зародыша содержатся большие темные бластомеры (макромеры) с базофильной цитоплазмой, делятся медленнее и образуют в совокупности эмбриобласт, или зачаток зародыша.
Зародыш, не имеет полости, называется морулы (от греч. Morula - ягода шелковицы). Хотя этот термин характерен для ранних зародышей хордовых (например, ланцетника, который в действительности напоминает на начальных этапах развития ягоду шелковицы), он употребляется также для обозначения ранних зародышей высших позвоночных, которые еще не имеют полости и представляют собой плотное скопление клеток.
Затем в зародыше появляется небольшая, заполненная жидкостью полость и он принимает вид шарика, который называется бластоцистой (бластулы). Стенка бластоцисты образована трофобластом. На внутренней поверхности зародыша расположен эмбриобласт.
В середине шестого дня в зародыше насчитывается более ста клеток. В этот период он находится в матке, где происходит его имплантация (от лат. Иnрlаntatio - проникновение), то есть углубление в слизистую оболочку. Количество бластомеров в зародыше при этом увеличивается.
В конце первой недели развития из трофобласта в процессе имплантации образуется двухслойный эпителий, внутренний слой которого называется цитотрофобласт. Он представлен клетками кубической формы. Снаружи находится плазмодиотрофобласт, который называют также синцитиотрофобласты, он представляет собой симпласты, т.е. неподеленную на клеточные территории цитоплазматическую массу с многочисленными ядрами.
Проникновение зародыша в слизистую оболочку матки связано с действием трофобласта, который выделяет гистолитические ферменты, обусловливающие локальное (местное) разрушение внутренней оболочки матки - эндометрия. Зародыш погружается в его толщу и эндометрий смыкается над ним. Таким образом, дальнейшее развитие эмбриона происходит в слизистой оболочке матки.
Трофобласт обеспечивает питание зародыша на ранних этапах его развития, активно участвует в имплантации зародыша и в формировании временного органа - хориона который, взаимодействуя со слизистой оболочкой матки, образует плаценту - внезародышевых орган, который обеспечивает связь зародыша с организмом матери. Развитие, строение и функциональное значение плаценты рассматриваются в курсах эмбриологии и гистологии.
С клеточных элементов эмбриобласта развивается зародыш и другие внезародышевые органы. Одна из существенных особенностей развития человека - очень раннее образование внезародышевой мезодермы. Источник ее возникновения - эмбриобласт. Из него выселяются клетки, в виде полосок заполняют полость бластоцист и выстилают изнутри трофобласт. Внезародышевая мезодерма быстро превращается в внезародышевую мезенхиму, в результате чего внешняя оболочка зародыша состоит из трофобластического эпителия и мезенхимы. Она называется хорионом. Как отмечалось выше, хорион образует зародышевую часть плаценты - органа, обеспечивающего связь зародыша с организмом матери.
Затем начинается гаструляция, которая делится на два этапа. На первом образуются зародышевые листки, а на втором - осевой комплекс, определяющий общий план строения тела позвоночных и человека и зародышевых зачатков, из которых образуются различные ткани.
Зародышевыми листками называют первичные пласты клеток, образующихся в первой фазе гаструляции. Они отличные по расположению (наружный листок - эктодерма, внутренний - энтодерма и средний - мезодерма), размерами и строением клеток и направленностью дальнейшего развития. К осевому комплексу относятся нервная трубка, хорда (спинная струна), дорсальная мезодерма, расположенная по бокам от нервной трубки и хорды, первичная кишка. Во второй фазе гаструляции образуются также эмбриональные зачатки - комплексы клеток, которые возникают из зародышевых листков и начинают разные ткани. Таким образом, из одного и того же эмбрионального листка образуются различные ткани.
Морфологическим проявлением первой фазы гаструляции, который наблюдается в 7,5-недельного зародыша, является разделение эмбриобласта на два слоя - внешний (эпибласт) и внутренний (гипобласт).
Эпибласт имеет зачатки эктодермы, зародышевой мезодермы, хорды (спинной струны) и зародышевой (кишечной) энтодермы. Гипобласт является зачатком внезародышевой или желтковой энтодермы, что представляет собой ту часть внутреннего зародышевого листка, которая впоследствии войдет в состав одного из временных органов - желточного мешка.
Эндодермальных клетки, отделившиеся от эпибласта, начинают обрастать вокруг полости, образовавшейся в расположенных ниже участках внезародышевой мезодермы. Постепенно они приобретают вид чаши, края которой срастаются, в результате чего образуется желточный пузырек. Этот процесс с началом образования желточного мешка аналогичный тому, который характеризует развитие этого внезародышевого органа у млекопитающих.
Впоследствии, после начала имплантации, в эпибласте появляется первичная амниотическая полость за счет появления жидкости, которая раздвигает клетки. Затем кровля этой полости (амниотического пузырька) раскрывается и временно отстраняется извне участком цитотрофобласта. После этого стенки амниотического пузырька растут вверх и сливаются, вследствие чего полость амниотического пузырька снова становится замкнутой, окруженной клетками эпибласта. Этот процесс напоминает возникновение так называемых амниотических складок у птиц и млекопитающих.
С 9 до 14 суток развития зародыш человека имеет такое строение: внешняя стенка, образованная хорионом, состоит из внезародышевой мезенхимы и трофобластического эпителия. Последний, как отмечалось ранее, включает в свой состав цитотрофобласт и плазмодиотрофобласт. В утолщенной стенке зародыша, которая обращена вглубь стенки матки, в внезародышевой мезенхиме содержатся два пузырька, которые контактируют между собой, - амниотический и желточный. Участок амниотического пузырька, прилегающий к желточному, образует утолщение - так называемый зародышевый щиток, из которого формируется тело зародыша. От мезенхимы, ограничивают два пузырьки, к хориону тянется полоска клеток - амниотическая ножка, место отхождения которой соответствует заднему концу тела зародыша.
Благодаря описанным выше процессам зародыш обеспечивается тремя внезародышевыми органами - хорионом, участвующий в образовании плаценты, амнионом и желточным мешком. Из большей части амниотического пузырька формируется внезародышевый орган - амнион, одной из функций которого является образование амниотической жидкости - искусственной водной среды для развивающегося зародыша. С гипобласта образуется желточный пузырек (мешок), который не содержится у человека и большинства млекопитающих желтка, но выполняет важную роль первого органа кроветворения, участвующий в развитии первичных половых клеток.
Таким образом, в процессе эмбрионального развития человека появление ряда внезародышевых органов (хориона, амниона и желточного мешка) предшествует возникновению тела зародыша, что является свидетельством первоочередного создания условий, необходимых для развития эмбриона.
На 15 сутки внутриутробного развития начинается вторая фаза гаструляции, которая происходит одинаково как у птиц, так и у плацентарных млекопитающих. Переход от первой фазы гаструляции ко второй осуществляется постепенно, объединяя признаки первой (образование зародышевых листков) и второй (формирование комплекса осевых органов). Внешним признаком второй фазы гаструляции является появление в заднем конце зародышевого щитка вытянутой в длину полоски клеток, которая называется первичной полоской. На переднем ее конце образуется небольшое поднятие клеток - первичный, или гензеновський узелок.
На вершине первичного узелка возникает маленькая углубление - первичная ямка. В центральной части первичной полоски появляется первичная бороздка, которая является продолжением первичного ямки. Часть материала наружного зародышевого листка через первичную ямку погружается внутрь и входит в состав переднего отдела энтодермы, образуя так называемую передхордальную пластинку, из которой образуется эпителиально выстеленный передний отдел пищеварительного канала и две передние пары сомиты (сегментированных участков дорсальной мезодермы).
Ориентация первичной полоски определяет расположение осевых органов и, таким образом, общий план строения тела позвоночных.
Часть клеток эпибласта мигрирует через первичную полоску и, внедрившись в подчиненную часть гипобласта, образует зачаток кишечной энтодермы, которая впоследствии принимает участие в развитии кишки. Вторая часть гипобласта, как отмечалось выше, идет на образование желтковой энтодермы - внутреннего слоя желточного мешка.
В результате миграции клеток из эпибласта через первичную полоску также возникает зародышевая мезодерма, которая расширяется в латеральных направлениях под эпибласт.
С образованием зародышевой мезодермы тесно связано происхождение спинной струны - хорды. Ее зачаток образуется в виде так называемого хордального (или главного) отростка - тяжа клеток, образовавшихся за счет зародышевой мезодермы, который растет вперед от первоначального (гензенивського) узелка между эпибластом и гипобластом. Хорда определяет местонахождение будущего позвоночного столба, поскольку вокруг нее формируются позвонки, и существенно влияет на развитие нервной системы. У высших млекопитающих остатки хорды сохраняются и в постнатальном периоде развития в составе студенистого ядра, nucleus pulposus, межпозвоночных дисков.
Верхний слой клеток зародышевого щитка после образования хорды превращается в эктодерму. Под влиянием хорды или хордомезодермы на эктодерму, содержащейся над ней, последняя утолщается и превращается в нервную пластинку, затем в удлиненный нервный клубок, а в дальнейшем - в нервную трубку, за счет которой образуется как центральная, так и периферическая нервная система.
В конце второй - начале третьей недели эмбрионального развития закладывается четвертый внезародышевый орган - аллантоис. Сначала он имеет вид выроста желточного мешка, слепо заканчивается и врастает в амниотическую ножку. Позднее (после образования кишки) аллантоис отходит от передней стенки заднего отдела кишечной трубки. Вслед за образованием нервной трубки слои мезодермы, лежащие по бокам от нее и от хорды, подлежат сегментации. Латеральные отделы мезодермы, которые позже оказываются в составе брюшной части зародыша и названные поэтому брюшной мезодермой, расщепляются на два слоя: наружный, контактирующий с эктодермой (Париетальный листок) и внутренний, соединенный с энтодермой (висцеральный листок). Сомиты соединены с вентральной мезодермой тонкими тяжами клеток, называемых промежуточной мезодермой. Затем сомиты делятся на три части: дорсолатеральную (дерматом), медиовентральную (склеротом) и промежуточную (миотом). С появлением сомит начинается сомитный период развития зародыша. На 21-й день в его составе обнаруживается 2-3 пары сомиты; на 23-й день - 10 пар; на 25-й день - 14 пар; на 27-й день - 25 пар; в конце 5 недели - 43-44 пары.
Промежутки между зародышевыми листками и основными органами заполняются зародышевой соединительной тканью - мезенхима, которая состоит из веретенообразных или ведротчастых клеток, контактирующих отростками и погружены в аморфное межклеточное вещество. Мезенхима образуется, главным образом, из мезодермы, однако в ее развитии участвуют и другие зародышевые листки, из которых эктодерма (М. Ф. Кащенко).
Когда висцеральный листок вентральной мезодермы вступает в контакт с кишечной энтодермой, тело зародыша начинает отграничиваться от внезародышевых органов туловищными складками. Этот процесс начинается с вентрального прогиба краев зародыша, который в результате приобретает дорсальную выпуклость. Затем изогнутые края зародыша в главной и каудальной частях начинают углубляться. Расширение этих отделяя складки приводит к образованию боковых и брюшных стенок зародыша, а также возникновение так называемого желточного стебелька, которое связывает его с желточным мешком. С заглублением туловищной складки кишечная энтодерма и висцеральный листок вентральной мезодермы сворачиваются в трубку, что приводит к образованию кишки. Между тем, как последняя включает в свой состав кишечную ентодерму, желточный мешок содержит желтковую ентодерму. Аллантоис, который ранее имел вид выпячивания желточного мешка, теперь отходит от задней кишки.
С разрастанием амниотической оболочки растет объем амниона, который приближается к хориону и срастается с ним. Амниотическая ножка при этом перемещается из каудального участка в вентральные стенки зародыша и соединяется с амнионом, образуя пупочный канатик (пуповину). Аллантоис, ранее проник в амниотическую ножку, частично включается в состав пуповины. В процессе этих преобразований желточный мешок удаляется от зародыша, а его связь с кишечником сохраняется благодаря трубчатой желтково-кишечной протоке. Последний наряду с аллантоисом также находится в составе пупочного канатика. Впоследствии аллантоис и желтково-кишечная протока, входящие в состав пуповины, редуцируются.
 Стадии эмбриогенеза I – оплодотворение и образование зиготы; II – дробление и образование бластулы (бластоцисты); III – гаструляция – образование зародышевых листков и комплекса осевых органов; IV – гистогенез и органогенез зародышевых и внезародышевых органов; V – системогенез.
Стадии эмбриогенеза I – оплодотворение и образование зиготы; II – дробление и образование бластулы (бластоцисты); III – гаструляция – образование зародышевых листков и комплекса осевых органов; IV – гистогенез и органогенез зародышевых и внезародышевых органов; V – системогенез.
 Пренатальный период начинается с момента оплодотворения и образования зиготы и продолжается 280 дней. Эмбриогенез подразделяется на: - начальный период (первая неделя) - эмбриональный период (со 2 -й по 9 нед.) - плодный период (с 8 -9 нед. до рождения).
Пренатальный период начинается с момента оплодотворения и образования зиготы и продолжается 280 дней. Эмбриогенез подразделяется на: - начальный период (первая неделя) - эмбриональный период (со 2 -й по 9 нед.) - плодный период (с 8 -9 нед. до рождения).

 ГАМЕТОГЕНЕЗ ✓ Первичные половые клетки гоноциты (гонобласты) впервые выявляются в стенке желточного мешка (в конце третьей недели жизни эмбриона человека). ✓ Гоноциты являются индифферентыми клетками: они не разделяются на мужские и женские клетки. ✓ Гоноциты мигрируют в закладки половых желез и дифференцируются в сперматогонии (в семеннике) или в овогонии (в яичнике). Это процесс начинается на 6 -ой недели жизни эмбриона.
ГАМЕТОГЕНЕЗ ✓ Первичные половые клетки гоноциты (гонобласты) впервые выявляются в стенке желточного мешка (в конце третьей недели жизни эмбриона человека). ✓ Гоноциты являются индифферентыми клетками: они не разделяются на мужские и женские клетки. ✓ Гоноциты мигрируют в закладки половых желез и дифференцируются в сперматогонии (в семеннике) или в овогонии (в яичнике). Это процесс начинается на 6 -ой недели жизни эмбриона.
 СПЕРМАТОГЕНЕЗ происходит в извитых канальцах семенника. В процессе сперматогенеза выделяют 4 фазы (периода): размножение (пролиферация) рост созревание формирование (спермиогенез).
СПЕРМАТОГЕНЕЗ происходит в извитых канальцах семенника. В процессе сперматогенеза выделяют 4 фазы (периода): размножение (пролиферация) рост созревание формирование (спермиогенез).
 I. Фаза размножения: Мужские половые клетки представлены диплоидными клетками – СПЕРМАТОГОНИЯМИ, которые делятся МИТОТИЧЕСКИ. Различают 3 типа сперматогоний (рис. 1): тёмные А сперматогонии – истинные стволовые клетки, устойчивы к действию вредных факторов, делятся редко; светлые А сперматогонии – полустволовые клетки, способны к частым митотическим делениям (результат деления: образуются две А- сперматогонии, либо одна А и одна В-). светлые В сперматогонии, коммитированные клетки – готовятся к мейозу и вступают в фазу роста.
I. Фаза размножения: Мужские половые клетки представлены диплоидными клетками – СПЕРМАТОГОНИЯМИ, которые делятся МИТОТИЧЕСКИ. Различают 3 типа сперматогоний (рис. 1): тёмные А сперматогонии – истинные стволовые клетки, устойчивы к действию вредных факторов, делятся редко; светлые А сперматогонии – полустволовые клетки, способны к частым митотическим делениям (результат деления: образуются две А- сперматогонии, либо одна А и одна В-). светлые В сперматогонии, коммитированные клетки – готовятся к мейозу и вступают в фазу роста.
 Участок стенки извитого канальца семенника 1 – миоидные клетки стенки; 2 – базальная мембрана; 3 - сперматогонии А; 4 – сперматогонии В; 5 – сперматоцит первого порядка; 6 – сперматоцит второго порядка; 7 – сперматиды на ранней стадии развития; 8 – сперматиды в конце развития; 9 – клетка Сертоли.
Участок стенки извитого канальца семенника 1 – миоидные клетки стенки; 2 – базальная мембрана; 3 - сперматогонии А; 4 – сперматогонии В; 5 – сперматоцит первого порядка; 6 – сперматоцит второго порядка; 7 – сперматиды на ранней стадии развития; 8 – сперматиды в конце развития; 9 – клетка Сертоли.
 II. Фаза роста: В сперматогонии увеличиваются в объёме (в 4 раза) и вступают в профазу первого мейотического деления, перемещаясь из базального слоя по направлению к просвету канальца → СПЕРМАТОЦИТЫ 1 -го порядка (диплоидные).
II. Фаза роста: В сперматогонии увеличиваются в объёме (в 4 раза) и вступают в профазу первого мейотического деления, перемещаясь из базального слоя по направлению к просвету канальца → СПЕРМАТОЦИТЫ 1 -го порядка (диплоидные).
 III. Фаза созревания: В результате 1 -го деления мейоза (редукционного) образуются СПЕРМАТОЦИТЫ 2 го порядка, которые быстро вступают во 2 -е деление мейоза (эквационное), образующиеся клетки называются СПЕРМАТИДЫ (гаплоидные). Начиная со сперматогоний типа В, дочерние клетки при делении не отделяются полностью друг от друга, а остаются связанными с помощью цитоплазматических мостиков, образуя синцитий. Лишь зрелые сперматозоиды отделяются от синцития.
III. Фаза созревания: В результате 1 -го деления мейоза (редукционного) образуются СПЕРМАТОЦИТЫ 2 го порядка, которые быстро вступают во 2 -е деление мейоза (эквационное), образующиеся клетки называются СПЕРМАТИДЫ (гаплоидные). Начиная со сперматогоний типа В, дочерние клетки при делении не отделяются полностью друг от друга, а остаются связанными с помощью цитоплазматических мостиков, образуя синцитий. Лишь зрелые сперматозоиды отделяются от синцития.

 IV. Фаза формирования (спермиогенез): около 50 суток В процессе формирования зрелых сперматозоидов из сперматид происходят: конденсация ядерного хроматина; конденсация хроматина приводит к образованию суперкомпактного ядра, что увеличивает мобильность сперматозоида и защищает геном от повреждения; образование акросомы из элементов комплекса Гольджи; акросома - специальный тип лизосомы, содержит гидролитические ферменты (такие как гиалуронидаза и т. п.) для разрушения оболочек яйцеклетки; центриоли перемещаются к противоположному полюсу ядра; от дистальной центриоли образуется осевая нить (аксонема) жгутика. митохондрии в виде спирали окружают часть аксонемы, образуя утолщенный участок – промежуточный отдел; митохондрии обеспечивают энергию для движения сперматозоида; цитоплазма редуцируется до минимума; после сброса излишек цитоплазмы, сперматозоиды отделяются от общей ассоциации клеток - синцития - и становятся свободными.
IV. Фаза формирования (спермиогенез): около 50 суток В процессе формирования зрелых сперматозоидов из сперматид происходят: конденсация ядерного хроматина; конденсация хроматина приводит к образованию суперкомпактного ядра, что увеличивает мобильность сперматозоида и защищает геном от повреждения; образование акросомы из элементов комплекса Гольджи; акросома - специальный тип лизосомы, содержит гидролитические ферменты (такие как гиалуронидаза и т. п.) для разрушения оболочек яйцеклетки; центриоли перемещаются к противоположному полюсу ядра; от дистальной центриоли образуется осевая нить (аксонема) жгутика. митохондрии в виде спирали окружают часть аксонемы, образуя утолщенный участок – промежуточный отдел; митохондрии обеспечивают энергию для движения сперматозоида; цитоплазма редуцируется до минимума; после сброса излишек цитоплазмы, сперматозоиды отделяются от общей ассоциации клеток - синцития - и становятся свободными.


 I. Фаза размножения: В отличие от сперматогенеза, фаза размножения в овогенезе протекает исключительно в эмбриональный период развития. Овогонии делятся путём митозов. Большинство образующихся дочерних клеток погибает (этот процесс называется атрезия), часть вступает в фазу роста (в профазу первого деления мейоза). Митотические деления овогоний прекращаются к 7 месяцу эмбриогенеза. При рождении все яйцеклетки в яичнике являются овоцитами первого порядка (первичными овоцитами).
I. Фаза размножения: В отличие от сперматогенеза, фаза размножения в овогенезе протекает исключительно в эмбриональный период развития. Овогонии делятся путём митозов. Большинство образующихся дочерних клеток погибает (этот процесс называется атрезия), часть вступает в фазу роста (в профазу первого деления мейоза). Митотические деления овогоний прекращаются к 7 месяцу эмбриогенеза. При рождении все яйцеклетки в яичнике являются овоцитами первого порядка (первичными овоцитами).
 II. Фаза роста: Фаза роста соответствует жизни первичного овоцита, находящегося в профазе первого деления мейоза, делится на два периода: малого (до полового созревания) и большого роста. К моменту рождения в яичнике насчитывается около 2 млн первичных овоцитов; к периоду полового созревания большинство из них погибает, остаётся около 400 тысяч яйцеклеток. Рост овоцитов происходит в фолликулах – структурах, в которых овоцит окружен эпителиальными фолликулярными клетками. В период малого роста имеется два типа фолликулов: примордиальные фолликулы (овоцит окружен одним слоем уплощенных фолликулярных клеток) и первичные фолликулы (появляется блестящая (прозрачная оболочка), фолликулярные клетки имеют кубическую или призматическую форму). Вступление овоцитов в период большого роста происходит только под воздействием гормонов (фолликуло-стимулирующего гормона); этому периоду оогенеза соответствуют растущие (вторичные и третичные) фолликулы. Период роста может продолжаться от 12 до 50 лет.
II. Фаза роста: Фаза роста соответствует жизни первичного овоцита, находящегося в профазе первого деления мейоза, делится на два периода: малого (до полового созревания) и большого роста. К моменту рождения в яичнике насчитывается около 2 млн первичных овоцитов; к периоду полового созревания большинство из них погибает, остаётся около 400 тысяч яйцеклеток. Рост овоцитов происходит в фолликулах – структурах, в которых овоцит окружен эпителиальными фолликулярными клетками. В период малого роста имеется два типа фолликулов: примордиальные фолликулы (овоцит окружен одним слоем уплощенных фолликулярных клеток) и первичные фолликулы (появляется блестящая (прозрачная оболочка), фолликулярные клетки имеют кубическую или призматическую форму). Вступление овоцитов в период большого роста происходит только под воздействием гормонов (фолликуло-стимулирующего гормона); этому периоду оогенеза соответствуют растущие (вторичные и третичные) фолликулы. Период роста может продолжаться от 12 до 50 лет.
 III. Фаза созревания Первое деление мейоза завершается только перед овуляцией (овуляция - разрыв зрелого третичного фолликула и выброс яйцеклетки из яичника в брюшную полость). В результате мейоза I образуется овоцит второго порядка (вторичный овоцит) – гаплоидная клетка. Овоцит второго порядка вступает во второе деление мейоза, но это деление блокируется на стадии метафазы и завершается только при условии оплодотворения. За один менструально-овариальный цикл (~ 28 дней) происходит овуляция обычно одной яйцеклетки. Таким образом, за репродуктивный период жизни женщины (~ 3040 лет) только около 450 яйцеклеток достигают зрелости – готовности к оплодотворению.
III. Фаза созревания Первое деление мейоза завершается только перед овуляцией (овуляция - разрыв зрелого третичного фолликула и выброс яйцеклетки из яичника в брюшную полость). В результате мейоза I образуется овоцит второго порядка (вторичный овоцит) – гаплоидная клетка. Овоцит второго порядка вступает во второе деление мейоза, но это деление блокируется на стадии метафазы и завершается только при условии оплодотворения. За один менструально-овариальный цикл (~ 28 дней) происходит овуляция обычно одной яйцеклетки. Таким образом, за репродуктивный период жизни женщины (~ 3040 лет) только около 450 яйцеклеток достигают зрелости – готовности к оплодотворению.
 Мейотическое деление в овогенезе отличается тем, что протекает с неравной цитотомией: в мейозе I хромосомы поровну распределяются между дочерними клетками, но только одна из клеток (вторичный овоцит) сохраняет почти всю цитоплазму; вторая клетка (первое полярное, или редукционное, направительное тельце) имеет очень мелкие размеры, небольшое ядро, минимум цитоплазмы.
Мейотическое деление в овогенезе отличается тем, что протекает с неравной цитотомией: в мейозе I хромосомы поровну распределяются между дочерними клетками, но только одна из клеток (вторичный овоцит) сохраняет почти всю цитоплазму; вторая клетка (первое полярное, или редукционное, направительное тельце) имеет очень мелкие размеры, небольшое ядро, минимум цитоплазмы.


 Яйцеклетка человека является вторично олиголецитальной и изолецитальной: в цитоплазме равномерно распределено относительно небольшое количество желтка; причём, в эволюции это вторично: впервые такой тип яйцеклетки встречается у ланцентника.
Яйцеклетка человека является вторично олиголецитальной и изолецитальной: в цитоплазме равномерно распределено относительно небольшое количество желтка; причём, в эволюции это вторично: впервые такой тип яйцеклетки встречается у ланцентника.
 Специфические структуры цитоплазмы яйцеклетки ЖЕЛТОЧНЫЕ ГРАНУЛЫ - В мембранных гранулах содержатся фосфо- и липопротеины - фосфовитин и липовителлин. Некоторые из этих веществ образуются в печени женщины, другие - непосредственно в ооците I. КОРТИКАЛЬНЫЕ ГРАНУЛЫ - Находясь под плазмолеммой, эти гранулы содержат ферменты, которые после оплодотворения участвуют в кортикальной реакции МУЛЬТИВЕЗИКУЛЯРНЫЕ ТЕЛЬЦА - Данные тельца появляются в результате переваривания фагоцитированных частиц
Специфические структуры цитоплазмы яйцеклетки ЖЕЛТОЧНЫЕ ГРАНУЛЫ - В мембранных гранулах содержатся фосфо- и липопротеины - фосфовитин и липовителлин. Некоторые из этих веществ образуются в печени женщины, другие - непосредственно в ооците I. КОРТИКАЛЬНЫЕ ГРАНУЛЫ - Находясь под плазмолеммой, эти гранулы содержат ферменты, которые после оплодотворения участвуют в кортикальной реакции МУЛЬТИВЕЗИКУЛЯРНЫЕ ТЕЛЬЦА - Данные тельца появляются в результате переваривания фагоцитированных частиц
 Особенности состава цитоплазмы яйцеклетки В цитоплазме - очень высокое содержание компонентов белоксинтезирующей системы (рибосом, т. РНК, м. РНК). ✓Отсутствуют центриоли - в связи с этим, способность к делениям восстанавливается только тогда, когда в клетку попадают центриоли сперматозоида. ✓На поверхности плазмолеммы имеются микроворсинки. ✓Яйцеклетку (точнее, ооцит II), как и предшествующие ей клетки, окружают оболочки: блестящая, или прозрачная (zona pellucida, или Zp), и зернистая, образованная фолликулярными клетками.
Особенности состава цитоплазмы яйцеклетки В цитоплазме - очень высокое содержание компонентов белоксинтезирующей системы (рибосом, т. РНК, м. РНК). ✓Отсутствуют центриоли - в связи с этим, способность к делениям восстанавливается только тогда, когда в клетку попадают центриоли сперматозоида. ✓На поверхности плазмолеммы имеются микроворсинки. ✓Яйцеклетку (точнее, ооцит II), как и предшествующие ей клетки, окружают оболочки: блестящая, или прозрачная (zona pellucida, или Zp), и зернистая, образованная фолликулярными клетками.
 Блестящая оболочка Zona pellucida: состоит из гликопротеинов разных видов (Zp 1, Zp 2, Zp 3) и гликозамингликанов. Гликопротеины фракции Zp 3 являются рецепторами для сперматозоидов, а гликопротеины фракции Zp 2 после кортикальной реакции препятствуют полиспермии. Компоненты блестящей оболочки синтезируются фолликулярными клетками. Зернистая оболочка: фолликулярные клетки зернистой оболочки представляют собой часть фолликулярного эпителия, имеют длинные отростки, пронизывающие блестящую оболочку. Лучистый венец: под лучистым венцом подразумевают только внутренние части фолликулярных клеток с отходящими от них отростками, либо всю совокупность зернистого слоя и блестящей оболочки. Редукционные тельца: где-то в составе оболочек или под ними находятся редукционные тельца - другие (помимо яйцеклетки) продукты двух делений мейоза.
Блестящая оболочка Zona pellucida: состоит из гликопротеинов разных видов (Zp 1, Zp 2, Zp 3) и гликозамингликанов. Гликопротеины фракции Zp 3 являются рецепторами для сперматозоидов, а гликопротеины фракции Zp 2 после кортикальной реакции препятствуют полиспермии. Компоненты блестящей оболочки синтезируются фолликулярными клетками. Зернистая оболочка: фолликулярные клетки зернистой оболочки представляют собой часть фолликулярного эпителия, имеют длинные отростки, пронизывающие блестящую оболочку. Лучистый венец: под лучистым венцом подразумевают только внутренние части фолликулярных клеток с отходящими от них отростками, либо всю совокупность зернистого слоя и блестящей оболочки. Редукционные тельца: где-то в составе оболочек или под ними находятся редукционные тельца - другие (помимо яйцеклетки) продукты двух делений мейоза.


 Головка Плазматическая мембрана: Содержит специальные белки, участвующие в таксисе сперсматозоида и связывании с яйцеклеткой эти белки (те, что участвуют в связывании с яйцеклеткой) имеют высокую видовую специфичность. Акросома: Акросома - это уплощённый мембранный мешочек, который, двойной шапочкой, покрывает ядро. Там содержатся литические ферменты (акрозин, гиалуронидаза и др.), разрушающие оболочки яйцеклетки. Ядро: резко уплотнено и содержит гаплоидный набор хромосом.
Головка Плазматическая мембрана: Содержит специальные белки, участвующие в таксисе сперсматозоида и связывании с яйцеклеткой эти белки (те, что участвуют в связывании с яйцеклеткой) имеют высокую видовую специфичность. Акросома: Акросома - это уплощённый мембранный мешочек, который, двойной шапочкой, покрывает ядро. Там содержатся литические ферменты (акрозин, гиалуронидаза и др.), разрушающие оболочки яйцеклетки. Ядро: резко уплотнено и содержит гаплоидный набор хромосом.
 Хвост Шейка или связующий отдел: Содержит 2 центриоли. От одной из них начинается аксонема, или осевая нить хвоста, образованная микротрубочками по схеме (9 х2) + 2. Промежуточная часть: В этой части вокруг аксонемы 9 наружных фибрилл, митохондриальная спиральная оболочка и плазмолемма. Главная или основная часть: Здесь вокруг аксонемы - фибриллярное влагалище (9 наружных фибрилл и волокнистая оболочка), а также плазмолемма. Концевая часть: В этой части вокруг аксонемы остаётся только плазмолемма.
Хвост Шейка или связующий отдел: Содержит 2 центриоли. От одной из них начинается аксонема, или осевая нить хвоста, образованная микротрубочками по схеме (9 х2) + 2. Промежуточная часть: В этой части вокруг аксонемы 9 наружных фибрилл, митохондриальная спиральная оболочка и плазмолемма. Главная или основная часть: Здесь вокруг аксонемы - фибриллярное влагалище (9 наружных фибрилл и волокнистая оболочка), а также плазмолемма. Концевая часть: В этой части вокруг аксонемы остаётся только плазмолемма.
 Эякулят мужчины имеет объём 2 -3 мл и содержит 200 -350 млн сперматозоидов У человека головка сперматозоида сильно уплощена.
Эякулят мужчины имеет объём 2 -3 мл и содержит 200 -350 млн сперматозоидов У человека головка сперматозоида сильно уплощена.
 Клеточный состав эякулята человека А- зрелые, Б- незрелые 1, 2 - типичный сперматозоид 3 -12 – атипичные сперматозоиды 6– 7 – аномалия формы головки и акросомы 8 -9 – аномалия жгутика 13– 18 – незрелые мужские половые клетки 19 – эпителиальные клетки 20 – 22 - лейкоциты
Клеточный состав эякулята человека А- зрелые, Б- незрелые 1, 2 - типичный сперматозоид 3 -12 – атипичные сперматозоиды 6– 7 – аномалия формы головки и акросомы 8 -9 – аномалия жгутика 13– 18 – незрелые мужские половые клетки 19 – эпителиальные клетки 20 – 22 - лейкоциты
 Сперматозоиды (в отличие от яйцеклеток) неоднородны по виду половой хромосомы, содержащейся в их ядре: у 50 % Сз имеется Х-хромосома, а у других 50 % Сз - Y-хромосома Пол ребёнка определяется "полом" сперматозоида:
Сперматозоиды (в отличие от яйцеклеток) неоднородны по виду половой хромосомы, содержащейся в их ядре: у 50 % Сз имеется Х-хромосома, а у других 50 % Сз - Y-хромосома Пол ребёнка определяется "полом" сперматозоида:
 Оплодотворение У человека - моноспермальный тип оплодотворения: только один сперматозоид может проникнуть в яйцеклетку Оптимальный срок для оплодотворения - первые 24 часа после овуляции (хотя ооцит II может сохранять способность к оплодотворению ещё некоторое время). Оплодотворение в норме происходит в ампулярной части маточной трубы
Оплодотворение У человека - моноспермальный тип оплодотворения: только один сперматозоид может проникнуть в яйцеклетку Оптимальный срок для оплодотворения - первые 24 часа после овуляции (хотя ооцит II может сохранять способность к оплодотворению ещё некоторое время). Оплодотворение в норме происходит в ампулярной части маточной трубы
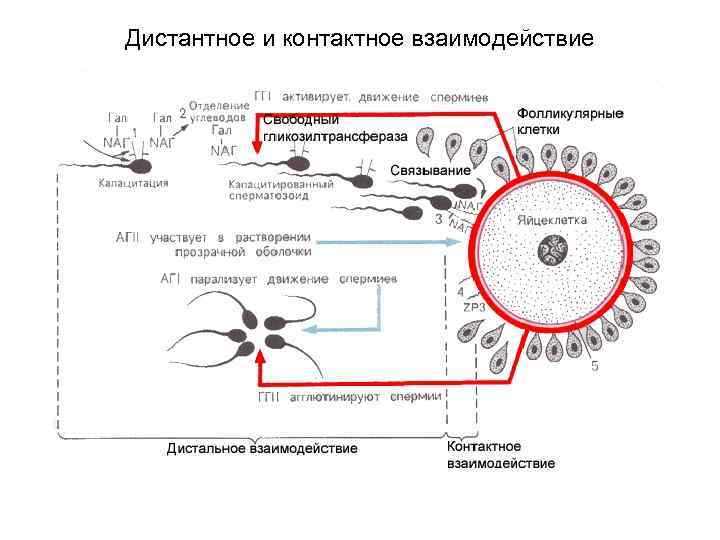
 Фазы оплодотворения 1. Дистантное взаимодействие и сближение гамет; 2. Контактное взаимодействие и активизация яйцеклетки; 3. Вхождение сперматозоида в яйцеклетку и последующее слияние – сингамия.
Фазы оплодотворения 1. Дистантное взаимодействие и сближение гамет; 2. Контактное взаимодействие и активизация яйцеклетки; 3. Вхождение сперматозоида в яйцеклетку и последующее слияние – сингамия.
 Оплодотворение 1, 2, 3, 4 – стадии акросомальной реакции 5 - блестящая зона 6 – перивителиновое пространство 7 – плазматическая мембрана 8 – кортикальная гранула 8 а – кортикальная реакция 9 – вхождение спермия в яйцеклетку 10 – зонная реакция
Оплодотворение 1, 2, 3, 4 – стадии акросомальной реакции 5 - блестящая зона 6 – перивителиновое пространство 7 – плазматическая мембрана 8 – кортикальная гранула 8 а – кортикальная реакция 9 – вхождение спермия в яйцеклетку 10 – зонная реакция
 I. Сближение и дистантное взаимодействие половых клеток Ооцит II медленно перемещается от яичника по направлению к матке; это происходит пассивно - благодаря току слизи. Данный ток вызывается биением ресничек мерцательных клеток и тоническими сокращениями маточных труб (под действием прогестерона). Во влагалище собственная подвижность сперматозоидов (Сз) невелика из-за имеющейся здесь кислой среды. В матку они попадают, в основном, пассивно - благодаря тоническим сокращениям женских половых путей. Затем часть Сз также, в основном, пассивно, достигает маточных труб. Капацитация т. е. активация Сз: метаболизм и подвижность Сз резко усиливаются, а мембраны Сз в области головки теряют поверхностные гликопротеины и поэтому приобретают способность связываться с блестящей оболочкой ооцита Капацитацию инициируют гиногамоны II, выделяемые ооцитом. Активное движение Сз (реотаксис, хемотаксис) Движение Сз становится преимущественно активным и обеспечивается биением их жгутиков. При этом сперматозоиды одновременно двигаются поступательно и вращаются вокруг своей оси;
I. Сближение и дистантное взаимодействие половых клеток Ооцит II медленно перемещается от яичника по направлению к матке; это происходит пассивно - благодаря току слизи. Данный ток вызывается биением ресничек мерцательных клеток и тоническими сокращениями маточных труб (под действием прогестерона). Во влагалище собственная подвижность сперматозоидов (Сз) невелика из-за имеющейся здесь кислой среды. В матку они попадают, в основном, пассивно - благодаря тоническим сокращениям женских половых путей. Затем часть Сз также, в основном, пассивно, достигает маточных труб. Капацитация т. е. активация Сз: метаболизм и подвижность Сз резко усиливаются, а мембраны Сз в области головки теряют поверхностные гликопротеины и поэтому приобретают способность связываться с блестящей оболочкой ооцита Капацитацию инициируют гиногамоны II, выделяемые ооцитом. Активное движение Сз (реотаксис, хемотаксис) Движение Сз становится преимущественно активным и обеспечивается биением их жгутиков. При этом сперматозоиды одновременно двигаются поступательно и вращаются вокруг своей оси;
 II. Контактное взаимодействие половых клеток Связывание клеток Достигая ооцита II, многочисленные Сз связываются с его оболочками (за счёт взаимодействия определённых рецепторов). При этом, из-за биения жгутиков Сз, ооцит начинает вращаться вокруг собственной оси. Акросомная реакция У связавшихся Сз развивается акросомная реакция: разрываются передние участки плазмолеммы и мембраны акросомы, отчего высвобождаются акросомальные ферменты: гиалуронидаза разъединяет клетки зернистой оболочки, а трипсиноподобный фермент акрозин и ряд других ферментов растворяют блестящую оболочку в месте прохождения Сз.
II. Контактное взаимодействие половых клеток Связывание клеток Достигая ооцита II, многочисленные Сз связываются с его оболочками (за счёт взаимодействия определённых рецепторов). При этом, из-за биения жгутиков Сз, ооцит начинает вращаться вокруг собственной оси. Акросомная реакция У связавшихся Сз развивается акросомная реакция: разрываются передние участки плазмолеммы и мембраны акросомы, отчего высвобождаются акросомальные ферменты: гиалуронидаза разъединяет клетки зернистой оболочки, а трипсиноподобный фермент акрозин и ряд других ферментов растворяют блестящую оболочку в месте прохождения Сз.
 III. Проникновение сперматозоида в ооцит В ооцит проникают ядро Сз и центриоли. После этого в ооците II в течение нескольких секунд развивается кортикальная реакция - выброс содержимого кортикальных гранул за пределы клетки, образуется оболочка оплодотворения. Кроме того, ооцитом выделяются гиногамоны I, которые вызывают агглютинацию оставшихся сперматозоидов. Завершение мейоза Одновременно проходят стадии второго деления мейоза (метафаза, анафаза, телофаза). Это увеличивает количество редукционных, или полярных, телец под блестящей оболочкой.
III. Проникновение сперматозоида в ооцит В ооцит проникают ядро Сз и центриоли. После этого в ооците II в течение нескольких секунд развивается кортикальная реакция - выброс содержимого кортикальных гранул за пределы клетки, образуется оболочка оплодотворения. Кроме того, ооцитом выделяются гиногамоны I, которые вызывают агглютинацию оставшихся сперматозоидов. Завершение мейоза Одновременно проходят стадии второго деления мейоза (метафаза, анафаза, телофаза). Это увеличивает количество редукционных, или полярных, телец под блестящей оболочкой.
 Подготовка зиготы к дроблению Сближение ядер В образующейся зиготе ядро Сз набухает (превращаясь в мужской пронуклеус и сближается с женским пронуклеусом (сближенные ядра называются синкарионом), но не сливается с ним. Удвоение ДНК и центриолей Удваиваются молекулы ДНК (в пронуклеусах) и пришедшие с Сз центриоли. Эти процессы продолжаются около суток.
Подготовка зиготы к дроблению Сближение ядер В образующейся зиготе ядро Сз набухает (превращаясь в мужской пронуклеус и сближается с женским пронуклеусом (сближенные ядра называются синкарионом), но не сливается с ним. Удвоение ДНК и центриолей Удваиваются молекулы ДНК (в пронуклеусах) и пришедшие с Сз центриоли. Эти процессы продолжаются около суток.
 Начало первого митотического деления Образование единой материнской звезды В первом митотическом делении участвуют два так и не слившихся пронуклеуса: их оболочки разрушаются, а хромосомы конденсируются и в метафазе образуют единую материнскую звезду. Всё это время продолжается медленное пассивное продвижение ооцита II, а затем зиготы, по маточной трубе к матке.
Начало первого митотического деления Образование единой материнской звезды В первом митотическом делении участвуют два так и не слившихся пронуклеуса: их оболочки разрушаются, а хромосомы конденсируются и в метафазе образуют единую материнскую звезду. Всё это время продолжается медленное пассивное продвижение ооцита II, а затем зиготы, по маточной трубе к матке.
 Рисунок - зигота человека на стадии синкариона. На рисунке вокруг зиготы видна плотная оболочка оплодотворения (1); она не имеет клеточной структуры, т. к. происходит из блестящей оболочки. Внутри зиготы - два ядра-пронуклеуса (2) почти равного объёма. Следовательно, очень мелкое и плотное ядро сперматозоида в результате деконденсации хромосом и набухания достигает размера ядра яйцеклетки. В каждом из этих ядер - гаплоидный набор хромосом, т. е. по 23 хромосомы. Поскольку ядра уже соприкасаются (что является сигналом к началу митоза), в них уже совершилось удвоение ДНК, и хромосомы стали двухроматидными
Рисунок - зигота человека на стадии синкариона. На рисунке вокруг зиготы видна плотная оболочка оплодотворения (1); она не имеет клеточной структуры, т. к. происходит из блестящей оболочки. Внутри зиготы - два ядра-пронуклеуса (2) почти равного объёма. Следовательно, очень мелкое и плотное ядро сперматозоида в результате деконденсации хромосом и набухания достигает размера ядра яйцеклетки. В каждом из этих ядер - гаплоидный набор хромосом, т. е. по 23 хромосомы. Поскольку ядра уже соприкасаются (что является сигналом к началу митоза), в них уже совершилось удвоение ДНК, и хромосомы стали двухроматидными
 Дробление Дробление зиготы– полное (голобластическое), неравномерное (2 типа бластомеров: мелкие, светлые и более крупные, тёмные), асинхронное, медленное. Плоскость первого дробления проходит через редукционные тельца.
Дробление Дробление зиготы– полное (голобластическое), неравномерное (2 типа бластомеров: мелкие, светлые и более крупные, тёмные), асинхронное, медленное. Плоскость первого дробления проходит через редукционные тельца.

 Дробление. Общая характеристика Сохранение оболочки оплодотворения На протяжении 2 -4 -х суток происходит дробление, т. е. совокупность митотических делений без периодов роста дочерних клеток. Рост клеток затруднён оттого, что вокруг зародыша сохраняется плотная оболочка оплодотворения, которая препятствует и притоку питательных веществ извне, (жизнедеятельность поддерживается за счёт расходования резервов яйцеклетки); и самому увеличению размера зародыша. Уменьшение размера клеток В силу вышесказанного, образуются всё более мелкие клетки и общий объём зародыша не увеличивается.
Дробление. Общая характеристика Сохранение оболочки оплодотворения На протяжении 2 -4 -х суток происходит дробление, т. е. совокупность митотических делений без периодов роста дочерних клеток. Рост клеток затруднён оттого, что вокруг зародыша сохраняется плотная оболочка оплодотворения, которая препятствует и притоку питательных веществ извне, (жизнедеятельность поддерживается за счёт расходования резервов яйцеклетки); и самому увеличению размера зародыша. Уменьшение размера клеток В силу вышесказанного, образуются всё более мелкие клетки и общий объём зародыша не увеличивается.

 Образование бластоцисты Через 4, 5 - 5 суток образуется бластоциста -зародышевый пузырёк, заполненный жидкостью. Компоненты бластоцисты следующие: Более мелкие светлые бластомеры образуют трофобласт. Трофобласт - однослойная стенка из мелких светлых клеток (впоследствии из трофобласта развивается внезародышевый орган - хорион). Более крупные темные – Эмбриобласт, или внутренняя клеточная масса- скопление крупных тёмных бластомеров в виде узелка на внутренней поверхности трофобласта у одного из полюсов. Бластоцель - полость, заполненная жидкостью.
Образование бластоцисты Через 4, 5 - 5 суток образуется бластоциста -зародышевый пузырёк, заполненный жидкостью. Компоненты бластоцисты следующие: Более мелкие светлые бластомеры образуют трофобласт. Трофобласт - однослойная стенка из мелких светлых клеток (впоследствии из трофобласта развивается внезародышевый орган - хорион). Более крупные темные – Эмбриобласт, или внутренняя клеточная масса- скопление крупных тёмных бластомеров в виде узелка на внутренней поверхности трофобласта у одного из полюсов. Бластоцель - полость, заполненная жидкостью.


 Рост бластоцисты Благодаря всасыванию трофобластом жидкости из полости матки, объём пузырька несколько увеличивается. В трофобласте появляются выросты, которые постепенно разрушают оболочку оплодотворения вокруг зародыша. В результате, за несколько часов до имплантации зародыш теряет эту оболочку. После этого оболочка уже не мешает зародышу увеличиваться в размере, и с этих пор митотические циклы клеток становятся обычными, т. е. включают фазу роста. Поэтому в последующем увеличение массы зародыша происходит гораздо быстрей.
Рост бластоцисты Благодаря всасыванию трофобластом жидкости из полости матки, объём пузырька несколько увеличивается. В трофобласте появляются выросты, которые постепенно разрушают оболочку оплодотворения вокруг зародыша. В результате, за несколько часов до имплантации зародыш теряет эту оболочку. После этого оболочка уже не мешает зародышу увеличиваться в размере, и с этих пор митотические циклы клеток становятся обычными, т. е. включают фазу роста. Поэтому в последующем увеличение массы зародыша происходит гораздо быстрей.
 Бластоциста остаётся свободной в полости матки до 67 дня э. р. , затем погружается в слизистый секрет маточных желез на поверхности эндометрия.
Бластоциста остаётся свободной в полости матки до 67 дня э. р. , затем погружается в слизистый секрет маточных желез на поверхности эндометрия.
 Имплантация – процесс проникновения зародыша в слизистую оболочку стенки матки (эндометрий) и установления тесных связей с её кровеносными сосудами – начало на 6 -7 день. Делится на две стадии - адгезию и инвазию Под действием маточного секрета блестящая оболочка растворяется, и бластоциста прикрепляется к эндометрию (адгезия). Клетки трофобласта быстро разрастаются, выделяют лизосомальные ферменты и разрушают ткани эндометрия. Обычно имплантация происходит в эндометрий задней стенки матки.
Имплантация – процесс проникновения зародыша в слизистую оболочку стенки матки (эндометрий) и установления тесных связей с её кровеносными сосудами – начало на 6 -7 день. Делится на две стадии - адгезию и инвазию Под действием маточного секрета блестящая оболочка растворяется, и бластоциста прикрепляется к эндометрию (адгезия). Клетки трофобласта быстро разрастаются, выделяют лизосомальные ферменты и разрушают ткани эндометрия. Обычно имплантация происходит в эндометрий задней стенки матки.

 Трофобласт С началом имплантации, (как только трофобласт входит в контакт с эпителием эндометрия), клетки трофобласта начинают активно делиться, дочерние клетки сливаются, образуя единую цитоплазматическую массу, содержащую множество ядер, и не имеющую клеточных границ, образуя симпласто- (синцитио)-трофобласт (наружный листок) - многоядерный симпласт, образующийся в результате слияния большого количества клеток. и цитотрофобласт - (внутренний листок) (сохраняет клеточное строение);
Трофобласт С началом имплантации, (как только трофобласт входит в контакт с эпителием эндометрия), клетки трофобласта начинают активно делиться, дочерние клетки сливаются, образуя единую цитоплазматическую массу, содержащую множество ядер, и не имеющую клеточных границ, образуя симпласто- (синцитио)-трофобласт (наружный листок) - многоядерный симпласт, образующийся в результате слияния большого количества клеток. и цитотрофобласт - (внутренний листок) (сохраняет клеточное строение);

 После имплантации симпластотрофобласт адсорбирует продукты распада тканей – гистотрофный тип питания (первые две недели). Таким образом, роль симпластотрофобласта заключается в том, что: благодаря действию лизосомальных ферментов обеспечивает инвазию зародыша в стенку матки; он обеспечивает питание, необходимое для роста зародыша в первые две недели жизни, абсорбируя продукты распада тканей матки (гистотрофный тип питания); выполняет эндокринную функцию (стб - место образования гормонов, включая ХГ (гормон, стимулирующий образование прогестерона жёлтым телом - определяется в крови или в моче женщин, начиная с 10 -го дня беременности, и является основой теста на беременность.).
После имплантации симпластотрофобласт адсорбирует продукты распада тканей – гистотрофный тип питания (первые две недели). Таким образом, роль симпластотрофобласта заключается в том, что: благодаря действию лизосомальных ферментов обеспечивает инвазию зародыша в стенку матки; он обеспечивает питание, необходимое для роста зародыша в первые две недели жизни, абсорбируя продукты распада тканей матки (гистотрофный тип питания); выполняет эндокринную функцию (стб - место образования гормонов, включая ХГ (гормон, стимулирующий образование прогестерона жёлтым телом - определяется в крови или в моче женщин, начиная с 10 -го дня беременности, и является основой теста на беременность.).

 После полного погружения зародыша дефект в эндометрии вначале заполяется свернувшейся кровью, затем зарастает соединительной тканью и к 12 -13 дню покрывается эпителием. Вокруг зародыша образуются лакуны, заполняющиеся материнской кровью. В клетках стромы эндометрия развивается т. н. децидуальная реакция: разрастаются сосуды, появляется отёчность, клетки увеличиваются в объёме и накапливают гликоген и липиды.
После полного погружения зародыша дефект в эндометрии вначале заполяется свернувшейся кровью, затем зарастает соединительной тканью и к 12 -13 дню покрывается эпителием. Вокруг зародыша образуются лакуны, заполняющиеся материнской кровью. В клетках стромы эндометрия развивается т. н. децидуальная реакция: разрастаются сосуды, появляется отёчность, клетки увеличиваются в объёме и накапливают гликоген и липиды.
 Гаструляция (2 я неделя) Гаструляция у человека проходит в две фазы. 1)расщепления (деламинация) на 7 -е сутки, - одновременно с имплантацией, образуется эпибласт и гипобласт; 2)иммиграции на 14 – 15 сутки, образуется первичная полоска, головной узелок. 3)Между этими двумя фазами идёт образование внезародышевых органов, необходимых для успешного развития зародыша.
Гаструляция (2 я неделя) Гаструляция у человека проходит в две фазы. 1)расщепления (деламинация) на 7 -е сутки, - одновременно с имплантацией, образуется эпибласт и гипобласт; 2)иммиграции на 14 – 15 сутки, образуется первичная полоска, головной узелок. 3)Между этими двумя фазами идёт образование внезародышевых органов, необходимых для успешного развития зародыша.
 Гаструляция 1 -я фаза – деламинация – на 7 день эмбриогенеза. Эмбриобласт расщепляется на эпибласт и гипобласт - будущая внезародышевая (желточная) энтодерма.
Гаструляция 1 -я фаза – деламинация – на 7 день эмбриогенеза. Эмбриобласт расщепляется на эпибласт и гипобласт - будущая внезародышевая (желточная) энтодерма.

 Мезодерма Из эпибласта выселяются клетки, дающие начало внезародышевой мезодерме. Внезародышевая мезодерма образует губчатую структуру, образованную тяжами клеток. Постепенно обрастает стенки амниотического и желточного пузырька, формируя амниотическую оболочку (изнутри выстлана внезародышевой эктодермой, снаружи – внезародышевой мезодермой) и желточный мешок (изнутри выстлан внезародышевой энтодермой, снаружи – внезародышевой мезодермой). При подрастании мезодермы к трофобласту формируется хориональная оболочка (внезародышевая мезодерма + цито- и симпластотрофобласт) и небольшие полости сливаются в единую полость хориона (синоним - внезародышевый целом).
Мезодерма Из эпибласта выселяются клетки, дающие начало внезародышевой мезодерме. Внезародышевая мезодерма образует губчатую структуру, образованную тяжами клеток. Постепенно обрастает стенки амниотического и желточного пузырька, формируя амниотическую оболочку (изнутри выстлана внезародышевой эктодермой, снаружи – внезародышевой мезодермой) и желточный мешок (изнутри выстлан внезародышевой энтодермой, снаружи – внезародышевой мезодермой). При подрастании мезодермы к трофобласту формируется хориональная оболочка (внезародышевая мезодерма + цито- и симпластотрофобласт) и небольшие полости сливаются в единую полость хориона (синоним - внезародышевый целом).


 Образование желточного мешка. Разрастание внезародышевой мезодермы Клетки гипобласта мигрируют и смыкаются, образуя стенку желточного пузырька. Эпибласт – дно амниотического пузырька; Гипобласт – крыша желточного пузырька. Внезародышевая мезодерма (первичная мезенхима) образует губчатую структуру.
Образование желточного мешка. Разрастание внезародышевой мезодермы Клетки гипобласта мигрируют и смыкаются, образуя стенку желточного пузырька. Эпибласт – дно амниотического пузырька; Гипобласт – крыша желточного пузырька. Внезародышевая мезодерма (первичная мезенхима) образует губчатую структуру.
 Зародыш связан с формирующимся хорионом через амниотическую ножку, образованную внезародышевой мезодермой. Внезародышевая мезодерма врастает в выросты трофобласта, и первичные ворсинки хориона становятся вторичными (на 12 -13 е сутки).
Зародыш связан с формирующимся хорионом через амниотическую ножку, образованную внезародышевой мезодермой. Внезародышевая мезодерма врастает в выросты трофобласта, и первичные ворсинки хориона становятся вторичными (на 12 -13 е сутки).
 1 - зародышевый эпибласт. 2 - амниотическая эктодерма. 3 - амниотическая полость (между двумя предыдущими листками). 4 - желточный пузырек (образуется из гипобласта). Поверхности, обрастаемые внезародышевой мезодермой: 5 А - наружная поверхность амниотического пузырька; 5 Б - наружная поверхность желточного мешка; 5 В - внутренняя поверхность трофобласта. 6 - хорион: это трофобласт вместе с подросшей к нему мезодермой. 7 - амниотическая ножка: образуется из внезародышевой мезодермы и связывает зародыш с хорионом.
1 - зародышевый эпибласт. 2 - амниотическая эктодерма. 3 - амниотическая полость (между двумя предыдущими листками). 4 - желточный пузырек (образуется из гипобласта). Поверхности, обрастаемые внезародышевой мезодермой: 5 А - наружная поверхность амниотического пузырька; 5 Б - наружная поверхность желточного мешка; 5 В - внутренняя поверхность трофобласта. 6 - хорион: это трофобласт вместе с подросшей к нему мезодермой. 7 - амниотическая ножка: образуется из внезародышевой мезодермы и связывает зародыш с хорионом.

 Вторую неделю эмбриогенеза называют периодом «двоек»: образуются два слоя – эпибласт и гипобласт, которые составляют зародышевый диск; развиваются два пузырька – амниотический и желточный; дифференцируются два слоя трофобласта – цито- и симпластотрофобласт. К концу же этой недели трофобласт образует ветвящиеся выпячивания - первичные ворсины, после чего называется уже хорионом (ворсинчатой оболочкой).
Вторую неделю эмбриогенеза называют периодом «двоек»: образуются два слоя – эпибласт и гипобласт, которые составляют зародышевый диск; развиваются два пузырька – амниотический и желточный; дифференцируются два слоя трофобласта – цито- и симпластотрофобласт. К концу же этой недели трофобласт образует ветвящиеся выпячивания - первичные ворсины, после чего называется уже хорионом (ворсинчатой оболочкой).

 Таким образом, в конце первой недели – начале второй недели пренатального развития происходят такие очень важные и совпадающие по времени их протекания процессы, как: имплантация; первая фаза гаструляции; появление внезародышевой мезодермы и развитие внезародышевых органов: амниона, желточного мешка, хориона. В связи с этим, конец первой недели жизни эмбриона относится к критическим (определяющим) периодом развития (7- 8 день э. р.)
Таким образом, в конце первой недели – начале второй недели пренатального развития происходят такие очень важные и совпадающие по времени их протекания процессы, как: имплантация; первая фаза гаструляции; появление внезародышевой мезодермы и развитие внезародышевых органов: амниона, желточного мешка, хориона. В связи с этим, конец первой недели жизни эмбриона относится к критическим (определяющим) периодом развития (7- 8 день э. р.)
 3 -я неделя развития Вторая стадия гаструляции – иммиграция происходит на 14 -15 й день эмбриогенеза.
3 -я неделя развития Вторая стадия гаструляции – иммиграция происходит на 14 -15 й день эмбриогенеза.

 Клетки, иммигрирующие в зоне гензеновского узелка, образуют хордомезодермальный отросток, растущий под эпибластом по направлению к головному концу по срединной линии зародыша. Клетки этого отростка дифференцируются в головную мезодерму и хорду. Роль хорды в процессе развития: ✴хорда - ось будущего позвоночника ✴хорда - первичный индуктор и организатор: под влиянием сигнальных молекул клеток хорды происходит дифференцировка первичной эктодермы в нейроэктодерму и образование нервной пластинки и нервной трубки. ✴nucleus pulposus
Клетки, иммигрирующие в зоне гензеновского узелка, образуют хордомезодермальный отросток, растущий под эпибластом по направлению к головному концу по срединной линии зародыша. Клетки этого отростка дифференцируются в головную мезодерму и хорду. Роль хорды в процессе развития: ✴хорда - ось будущего позвоночника ✴хорда - первичный индуктор и организатор: под влиянием сигнальных молекул клеток хорды происходит дифференцировка первичной эктодермы в нейроэктодерму и образование нервной пластинки и нервной трубки. ✴nucleus pulposus


 Мезенхима Клетки мезенхимы выселяются из всех зародышевых листков, наиболее активно из висцерального листка спланхнотома. Из мезенхимы образуются нескольких типов тканей и структур: кровь; все типы соединительной ткани; гладкая мышечная ткань, кровеносные сосуды и эндокард сердца; микроглия центральной нервной системы.
Мезенхима Клетки мезенхимы выселяются из всех зародышевых листков, наиболее активно из висцерального листка спланхнотома. Из мезенхимы образуются нескольких типов тканей и структур: кровь; все типы соединительной ткани; гладкая мышечная ткань, кровеносные сосуды и эндокард сердца; микроглия центральной нервной системы.


 Третичные ворсинки хориона Кровеносные сосуды плода врастают в хориональные ворсинки, которые становятся третичными (появляются на 3 -й неделе развития).
Третичные ворсинки хориона Кровеносные сосуды плода врастают в хориональные ворсинки, которые становятся третичными (появляются на 3 -й неделе развития).

 Туловищная складка С 20 -го дня – начало обособления тела зародыша от внезародышевых оболочек - образуется туловищная складка. В результате образования туловищных складок тело зародыша приподнимается над провизорными органами и отделяется от них. При этом зародыш как бы скручивается в трубку. Одновременно это приводит к образованию из кишечной энтодермы кишечной трубки, которая отделяется от внезародышевой энтодермы желточного мешка.
Туловищная складка С 20 -го дня – начало обособления тела зародыша от внезародышевых оболочек - образуется туловищная складка. В результате образования туловищных складок тело зародыша приподнимается над провизорными органами и отделяется от них. При этом зародыш как бы скручивается в трубку. Одновременно это приводит к образованию из кишечной энтодермы кишечной трубки, которая отделяется от внезародышевой энтодермы желточного мешка.

 Амнион Образует замкнутую полость вокруг зародыша. Функции: создание водной среды определенного химического состава и давления для свободного развития, защита от механических, гравитационных стрессов. Постепенно полость разрастается. К 7 -й неделе развития амниотическая мезодерма входит в контакт с мезодермой хориона (амнио-хориональная оболочка). Кроме того, амниотический эпителий обрастает амниотическую ножку.
Амнион Образует замкнутую полость вокруг зародыша. Функции: создание водной среды определенного химического состава и давления для свободного развития, защита от механических, гравитационных стрессов. Постепенно полость разрастается. К 7 -й неделе развития амниотическая мезодерма входит в контакт с мезодермой хориона (амнио-хориональная оболочка). Кроме того, амниотический эпителий обрастает амниотическую ножку.

 Амнион функционирует до момента рождения (плодный пузырь). К концу беременности заполнен 1 -1, 5 л амниотической жидкости (околоплодные воды).
Амнион функционирует до момента рождения (плодный пузырь). К концу беременности заполнен 1 -1, 5 л амниотической жидкости (околоплодные воды).
 Желточный мешок Функционирует только на ранних стадиях развития: локализация первых очагов кроветворения и образования кровеносных сосудов, и первичных половых клеток. После 7 -8 недели подвергается регрессии, остаётся в виде тяжа клеток в пупочном канатике, направляющего кровеносные сосуды к плаценте. В отличие от птиц, у человека (и других млекопитающих) желточный мешок практически не содержит желтка, а заполнен серозной жидкостью.
Желточный мешок Функционирует только на ранних стадиях развития: локализация первых очагов кроветворения и образования кровеносных сосудов, и первичных половых клеток. После 7 -8 недели подвергается регрессии, остаётся в виде тяжа клеток в пупочном канатике, направляющего кровеносные сосуды к плаценте. В отличие от птиц, у человека (и других млекопитающих) желточный мешок практически не содержит желтка, а заполнен серозной жидкостью.
 Аллантоис развивается на 17 -18 -е сутки в виде небольшого слепого выроста вентральной стенки задней кишки. Аллантоис врастает в амниотическую ножку, в его стенке развиваются пупочные кровеносные сосуды, которые он подводит к хориону (проводник сосудов). На 2 -м месяце э. р. редуцируется и вместе с остатками желточного мешка образует клеточный тяж в составе пупочного канатика.
Аллантоис развивается на 17 -18 -е сутки в виде небольшого слепого выроста вентральной стенки задней кишки. Аллантоис врастает в амниотическую ножку, в его стенке развиваются пупочные кровеносные сосуды, которые он подводит к хориону (проводник сосудов). На 2 -м месяце э. р. редуцируется и вместе с остатками желточного мешка образует клеточный тяж в составе пупочного канатика.
 Хорион – плодная часть плаценты В формировании хориона различают три периода: предворсинчатый (7 -8 -е сутки), период образования ворсинок (до 50 -х суток), период котиледонов (с 50 по 90 -е сутки). Образован хориональной пластиной (внезародышевая мезодерма) и выростами пластины – ветвящимися третичными ворсинками, покрытые трофобластом.
Хорион – плодная часть плаценты В формировании хориона различают три периода: предворсинчатый (7 -8 -е сутки), период образования ворсинок (до 50 -х суток), период котиледонов (с 50 по 90 -е сутки). Образован хориональной пластиной (внезародышевая мезодерма) и выростами пластины – ветвящимися третичными ворсинками, покрытые трофобластом.
 Формирование первичных ворсин (конец 2 -й недели) К середине 2 -й недели развития зародыш уже полностью погружён в эндометрий, а трофобласт состоит из двух слоёв - наружного (симпластотрофобласта) и внутреннего (цитотрофобласта). К концу же этой недели трофобласт образует ветвящиеся выпячивания - первичные ворсины, после чего называется уже хорионом (ворсинчатой оболочкой).
Формирование первичных ворсин (конец 2 -й недели) К середине 2 -й недели развития зародыш уже полностью погружён в эндометрий, а трофобласт состоит из двух слоёв - наружного (симпластотрофобласта) и внутреннего (цитотрофобласта). К концу же этой недели трофобласт образует ветвящиеся выпячивания - первичные ворсины, после чего называется уже хорионом (ворсинчатой оболочкой).
 Формирование вторичных ворсин (начало 3 -й недели) В начале 3 -й недели в ворсины хориона проникает внезародышевая мезенхима, (обрастающая изнутри все поверхности, в т. ч. внутреннюю поверхность трофобластахориона). Тем самым формируется соединительнотканная строма ворсин, а сами ворсины становятся вторичными.
Формирование вторичных ворсин (начало 3 -й недели) В начале 3 -й недели в ворсины хориона проникает внезародышевая мезенхима, (обрастающая изнутри все поверхности, в т. ч. внутреннюю поверхность трофобластахориона). Тем самым формируется соединительнотканная строма ворсин, а сами ворсины становятся вторичными.
 Формирование третичных ворсин (конец3 -й недели) К концу 3 -й недели внутри ворсин из клеток мезенхимы формируются первичные кровеносные сосуды, которые построены как гемокапилляры и вступают в связь с сосудами мезодермальной пластинки хориона. Ворсины с развитыми кровеносными сосудами называются третичными.
Формирование третичных ворсин (конец3 -й недели) К концу 3 -й недели внутри ворсин из клеток мезенхимы формируются первичные кровеносные сосуды, которые построены как гемокапилляры и вступают в связь с сосудами мезодермальной пластинки хориона. Ворсины с развитыми кровеносными сосудами называются третичными.
 Часть хориона, разрушающая стенку матки и участвующая в образовании плаценты, формирует сложноразветвленные ворсинки и носит название ворсинчатый хорион. Остальную поверхность составляет гладкий хорион. Самые крупные ворсинки, отходящие от хориональной пластины, носят название стволовых ворсинок. Стволовые ворсинки обильно ветвятся, самые мелкие веточки носят названия терминальных ворсинок. Ворсинки, внедряющиеся в базальную пластинку эндометрия, называются якорными ворсинками. Обычно стволовые ворсинки являются якорными.
Часть хориона, разрушающая стенку матки и участвующая в образовании плаценты, формирует сложноразветвленные ворсинки и носит название ворсинчатый хорион. Остальную поверхность составляет гладкий хорион. Самые крупные ворсинки, отходящие от хориональной пластины, носят название стволовых ворсинок. Стволовые ворсинки обильно ветвятся, самые мелкие веточки носят названия терминальных ворсинок. Ворсинки, внедряющиеся в базальную пластинку эндометрия, называются якорными ворсинками. Обычно стволовые ворсинки являются якорными.

 Плацента человека Плацента – единственный орган, состоящий из клеток двух генетически различных организмов. Состоит из плодной части (хорион с ворсинками) материнской части - измененный эндометрий
Плацента человека Плацента – единственный орган, состоящий из клеток двух генетически различных организмов. Состоит из плодной части (хорион с ворсинками) материнской части - измененный эндометрий
 Плацентация – период эмбриогенеза, на протяжении которого происходит развитие плаценты, один из критических периодов эмбриогенеза, соответствует 3 -8 неделям беременности. Поверхностный слой эндометрия - получил название децидуальной (отпадающей) оболочки. В зависимости от расположения относительно места имплантации различают: Decidua parietalis (пристеночная) – эндометрий, выстилающий полость матки за исключением участка имплантации; Decidua capsularis (сумочная) – часть эндометрия, которая окружает развивающийся эмбрион, образуя поверх него капсулу, и отделяет зародыш от полости матки (до 16 -й нед.) Decidua basalis (основная), материнская часть плаценты, та часть эндометрия, которая находится между плодом и базальным слоем эндометрия. Итак, материнская часть плаценты представлена: базальной пластинкой (decidua basalis) эндометрия; лакунами, заполненными материнской кровью.
Плацентация – период эмбриогенеза, на протяжении которого происходит развитие плаценты, один из критических периодов эмбриогенеза, соответствует 3 -8 неделям беременности. Поверхностный слой эндометрия - получил название децидуальной (отпадающей) оболочки. В зависимости от расположения относительно места имплантации различают: Decidua parietalis (пристеночная) – эндометрий, выстилающий полость матки за исключением участка имплантации; Decidua capsularis (сумочная) – часть эндометрия, которая окружает развивающийся эмбрион, образуя поверх него капсулу, и отделяет зародыш от полости матки (до 16 -й нед.) Decidua basalis (основная), материнская часть плаценты, та часть эндометрия, которая находится между плодом и базальным слоем эндометрия. Итак, материнская часть плаценты представлена: базальной пластинкой (decidua basalis) эндометрия; лакунами, заполненными материнской кровью.
 Kотиледоны Ворсинки, обращенные к decidua basalis, распределены неравномерно, группами – котиледонами. Котиледон – структурно-функциональная единица сформированной плаценты. Он образован стволовой ворсинкой и её разветвлениями. Котиледоны частично разделены соединительнотканными септами (плацентарными перегородками).
Kотиледоны Ворсинки, обращенные к decidua basalis, распределены неравномерно, группами – котиледонами. Котиледон – структурно-функциональная единица сформированной плаценты. Он образован стволовой ворсинкой и её разветвлениями. Котиледоны частично разделены соединительнотканными септами (плацентарными перегородками).

 Гематоплацентарный барьер Кровь матери и ребенка никогда не смешивается Компоненты гематоплацентарного барьера, разделяющего кровь матери и кровь плода: эндотелий капилляра плода; базальная мембрана в стенке капилляров плода; соединительная ткань ворсинок (с клеткамимакрофагами); базальная мембрана трофобласта; цитоторофобласт; симпластотрофобласт. Связь между циркуляциями крови плода и матери осуществляется через пупочный канатик
Гематоплацентарный барьер Кровь матери и ребенка никогда не смешивается Компоненты гематоплацентарного барьера, разделяющего кровь матери и кровь плода: эндотелий капилляра плода; базальная мембрана в стенке капилляров плода; соединительная ткань ворсинок (с клеткамимакрофагами); базальная мембрана трофобласта; цитоторофобласт; симпластотрофобласт. Связь между циркуляциями крови плода и матери осуществляется через пупочный канатик
 Гематоплацентарный барьер Кровь матери (1) и ребенка (8) не смешивается: - эндотелий капилляров плода (7); - базальная мембрана в стенке капилляров плода (6); - соединительная ткань ворсинок (5); - базальная мембрана трофобласта (4); - цитоторофобласт (3); - симпластотрофобласт (2)
Гематоплацентарный барьер Кровь матери (1) и ребенка (8) не смешивается: - эндотелий капилляров плода (7); - базальная мембрана в стенке капилляров плода (6); - соединительная ткань ворсинок (5); - базальная мембрана трофобласта (4); - цитоторофобласт (3); - симпластотрофобласт (2)

 Гематоплацентарный проницаем для ряда веществ и возбудителей болезней Возбудители болезней Вирусы (ВИЧ, цитомегаловирус, краснухи, оспы, ветряной оспы, кори, полиомиелита) Бактерии (туберкулез, Treponema) Простейшие (Toxoplasma) Наркотические и лекарственные средства Кокаин, алкоголь, кофеин, никотин, зоокумарин, дифенин тетрациклин противораковые препараты, анестетики, успокаивающие, анальгетики Антитела к резус-фактору
Гематоплацентарный проницаем для ряда веществ и возбудителей болезней Возбудители болезней Вирусы (ВИЧ, цитомегаловирус, краснухи, оспы, ветряной оспы, кори, полиомиелита) Бактерии (туберкулез, Treponema) Простейшие (Toxoplasma) Наркотические и лекарственные средства Кокаин, алкоголь, кофеин, никотин, зоокумарин, дифенин тетрациклин противораковые препараты, анестетики, успокаивающие, анальгетики Антитела к резус-фактору
 Функции плаценты трофическая (из организма матери к плоду поступают самые разнообразные питательные вещества, электролиты, витамины, некоторые гормоны; дыхательная (транспорт кислорода в кровь плода и перенос углекислого газа в кровь матери); выделительная (из крови плода в кровь матери поступают продукты метаболизма и выделяются через почки матери); защитная (препятствует развитию иммунного конфликта, проникновению лекарственных веществ – барьер не абсолютный); эндокринная (синтез ряда гормонов и других биологически активных веществ).
Функции плаценты трофическая (из организма матери к плоду поступают самые разнообразные питательные вещества, электролиты, витамины, некоторые гормоны; дыхательная (транспорт кислорода в кровь плода и перенос углекислого газа в кровь матери); выделительная (из крови плода в кровь матери поступают продукты метаболизма и выделяются через почки матери); защитная (препятствует развитию иммунного конфликта, проникновению лекарственных веществ – барьер не абсолютный); эндокринная (синтез ряда гормонов и других биологически активных веществ).
 Обмен через плацентарный барьер Из материнской крови в кровь плода Из крови плода в материнскую (снабжение) кровь (выделение) кислород питательные вещества (глюкоза, аминокислоты, свободные жирные кислоты, витамины) электролиты вода стероидные гормоны (кортизол, эстроген) антитела (Ig. G – обеспечивают пассивный иммунитет плода) белки плазмы крови (альбумин) некоторые белковые гормоны (тироксин, инсулин) эритроциты (очень мало) углекислый газ метаболиты (мочевина, мочевая кислота, билирубин, креатин, креатинин) стероидные гормоны (хорионический гонадотропин, плацентарный прогестерон) вода эритроциты (очень мало)
Обмен через плацентарный барьер Из материнской крови в кровь плода Из крови плода в материнскую (снабжение) кровь (выделение) кислород питательные вещества (глюкоза, аминокислоты, свободные жирные кислоты, витамины) электролиты вода стероидные гормоны (кортизол, эстроген) антитела (Ig. G – обеспечивают пассивный иммунитет плода) белки плазмы крови (альбумин) некоторые белковые гормоны (тироксин, инсулин) эритроциты (очень мало) углекислый газ метаболиты (мочевина, мочевая кислота, билирубин, креатин, креатинин) стероидные гормоны (хорионический гонадотропин, плацентарный прогестерон) вода эритроциты (очень мало)
 Типы плацент у млекопитающих Эпителиохориальная (свиньи): ворсинки хориона проникают в углубления слизистой оболочки (маточные железы), не разрушая их. Десмохориальная плацента (копытные): ворсинки хориона контактируют с соединительной тканью эндометрия матери. Эндотелио- (вазо-)хориальная (хищники) плацента: ворсинки хориона контактируют со стенкой сосудов эндометрия матери, не разрушая стенки. Гемохориальная плацента: ворсинки хориона разрушают эпителий, соединительную ткань, стенки сосудов матери и омываются кровью, заполняющие лакуны.
Типы плацент у млекопитающих Эпителиохориальная (свиньи): ворсинки хориона проникают в углубления слизистой оболочки (маточные железы), не разрушая их. Десмохориальная плацента (копытные): ворсинки хориона контактируют с соединительной тканью эндометрия матери. Эндотелио- (вазо-)хориальная (хищники) плацента: ворсинки хориона контактируют со стенкой сосудов эндометрия матери, не разрушая стенки. Гемохориальная плацента: ворсинки хориона разрушают эпителий, соединительную ткань, стенки сосудов матери и омываются кровью, заполняющие лакуны.
Оплодотворение
Оплодотворением называется процесс слияния зрелых мужской (сперматозоид) и женской (яйцеклетка) половых клеток, в результате чего возникает зигота, несущая генетическую информацию как отца, так и матери.
Процесс созревания мужских и женских половых клеток очень сложен. Сперматогенез совершается в извитых семенных канальцах мужских гонад. Он завершается в период половой зрелости образованием зрелых сперматозоидов, обладающих способностью к оплодотворению. Полному созреванию предшествует процесс редукционного деления, в результате которого в ядре сперматозоида содержится гаплоидный набор хромосом.
Сперматозоиды бывают двух видов: носители половых Х- и Y-хромосом. При слиянии с яйцеклеткой сперматозоида, являющегося носителем половой Х-хромо-сомы, из образующейся зиготы развивается эмбрион женского пола, при слиянии сперматозоида, имеющего половую Y-хромосому, возникает зародыш мужского пола (яйцеклетка всегда является носительницей половой Х-хромосомы).
Зрелый сперматозоид имеет длину до 50-60 мкм и состоит из головки, шейки и хвостовой части. Головка сперматозоида, имеющая овальную форму, содержит ядро, окруженное тонким слоем протоплазмы. Шейка имеет протоплазму, содержащую видоизмененную центросому, которая способствует процессу дробления оплодотворенной яйцеклетки. Хвостик состоит из протоплазмы и выполняет двигательные функции. В результате колебательных движений хвостовой части сперматозоиды способны совершать самостоятельные движения в половом тракте женщины со скоростью 2-3 мм/мин. Способность к движениям сперматозоиды получают после их контакта с секретом семенных пузырьков и предстательной железы. Такая смесь носит название семенной жидкости, или спермы. При половом сношении во влагалище женщины изливается в среднем около 3-5 мл спермы, в которой содержится 300-500 млн сперматозоидов. Использование электронной микроскопии позволило установить весьма сложную структуру сперматозоида (рис. 3.1).
Семенная жидкость имеет сложный состав и содержит фруктозу, белковые вещества, протеазы, кислую фосфатазу, лимонную кислоту и биологически активные вещества - простагландины. Семенная жидкость обладает выраженной антигенной активностью, при этом антигены обнаружены как в самих сперматозоидах, так и в жидкой части спермы. Полагают, что число антигенных компонентов в семенной жидкости достигает 12. В процессе оплодотворения происходит нейтрализация антигенов сперматозоидов.
Развитие яйцеклеток связано с ростом и развитием первичных фолликулов, находящихся в корковом слое яичников. Созревание яйцеклетки условно можно разделить на созревание ядра и созревание цитоплазмы. Под созреванием ядра понимают совокупность процессов, переводящих ядро из стадии диплотена I (или стадии герминативного пузырька) до метафазы второго мейотического деления. Созревание ядра не включает в себя завершение мейоза, так как только проникновение сперматозоида внутрь яйцеклетки вызывает завершение второго редукционного деления.
Головка Тело Хвост
Рис.3.1. Строение сперматозоида человека (электронно-микроскопическая схема). 1 - акросома; 2 - ядро; 3 - шейка; 4 - митохондрии; 5 - осевые нити.

Рис. 3.2. Зрелая яйцеклетка, окруженная клетками лучистого венца.
От периода наступления полового созревания до менопаузы у женщины в каждом менструальном цикле обычно созревает одна яйцеклетка. Овоцит первого порядка превращается в овоцит второго порядка при отщеплении первого полярного тельца. В момент овуляции овоцит второго порядка оказывается блокированным на стадии мета-фазы второго мейотического деления. Созревание яйцеклеток стало известно в деталях благодаря возможности культивирования яйцеклеток in vitro, что в настоящее время широко используют при экстракорпоральном оплодотворении.
Зрелая яйцеклетка состоит из ядра, цитоплазмы, окружена блестящей оболочкой и клетками лучистого венца, представляющего собой остатки гранулезных клеток фолликула (рис. 3.2). Женская половая клетка, как и мужская, обладает антигенными свойствами. Особенно богата различными антигенами ее блестящая оболочка.
Прежде чем анализировать механизмы оплодотворения, т.е. процесс слияния яйцеклетки со сперматозоидом, следует осветить вопросы транспорта гамет. В первую очередь это касается продвижения сперматозоидов по каналам репродуктивной системы женщины. При нормальной эякуляции во влагалище попадает в среднем около 100 млн сперматозоидов, некоторые из которых имеют те или иные морфологические либо функциональные отклонения. Часть сперматозоидов, в том числе и неполноценных, остается во влагалище и подвергается фагоцитозу. Вместе со сперматозоидами во влагалище попадают и другие составные части спермы, при этом особая роль принадлежит простагландинам. Под их влиянием происходит активация сократительной активности матки и маточных труб, что очень важно для нормального транспорта гамет.
Из влагалища, имеющего у здоровой женщины кислую среду (кислая
среда неблагоприятна для жизнедеятельности сперматозоидов), последние быстро поступают в цервикальную слизь, которая во время полового акта под влиянием сокращений мышц шейки матки выделяется из цервикального канала. Наличие слабощелочной реакции цервикальной слизи способствует повышению двигательной активности сперматозоидов.
Оптимальный состав цервикальной слизи формируется к моменту овуляции в основном под влиянием эстрогенных гормонов яичников. В этот период мицелии слизи располагаются в виде своеобразных цепочек, ориентированных по силовым линиям магнитного поля земли. Сперматозоиды продвигаются по мицелиям слизи по направлению к матке, но лишь при определенной архитектонике цепочек мицелл. Турбулентные движения сперматозоидов наиболее выражены в пристеночных областях шейки матки, при этом часть сперматозоидов на некоторое время может депонироваться в криптах шейки матки, создавая тем самым своеобразный резерв спермы, откуда в дальнейшем может происходить дополнительное их поступление вверх по каналам репродуктивной системы.
В верхних отделах полового тракта женщины начинается процесс, называемый капацитацией спермы, - приобретение ею благодаря сложным изменениям способности к оплодотворению. В результате капацитации сперматозоиды приобретают способность к акросомальной реакции. Помимо этого, капацитация выражается в изменениях движений хвостовых частей сперматозоидов (наличие сверхактивной подвижности). Тонкие механизмы капацитации до настоящего времени полностью не изучены. Время капацитации различно у разных сперматозоидов, что, по-видимому, является важной приспособительной реакцией для процесса оплодотворения. Капацити-рованные сперматозоиды очень активны, однако продолжительность их жизни меньше, чем некапацитированных. Капацитированные сперматозоиды обладают повышенной способностью пенетрировать ткани, что имеет решающее значение в процессе оплодотворения яйцеклетки.
Транспорт сперматозоидов в матку, а затем и в маточные трубы в основном обеспечивается сокращениями гладкой мускулатуры этих органов. Полагают также, что трубно-маточные сфинктеры являются своеобразными дозаторами поступления сперматозоидов из полости матки в просветы маточных труб.
Наряду с сократительной способностью маточных труб, которые находятся под сложным гормональным воздействием (эстрогены, андрогены, окситоцин), а также под влиянием простагландинов, большое значение в продвижении сперматозоидов, помимо их собственной высокой кинетической активности, принадлежит таким факторам, как движения микроворсин реснитчатого эпителия эндоцервикса и ток жидкости в просвете маточной трубы. Таким образом, перемещение сперматозоидов по каналам репродуктивной системы женщины представляет собой чрезвычайно сложный многокомпонентный процесс.
Результаты современных научных исследований свидетельствуют о наличии двухфазности процесса транспорта сперматозоидов по маточным трубам. В первую (короткую) фазу сперматозоиды начинают быстро поступать в ампулу трубы. Эта фаза длится всего несколько минут и регулируется сократительной активностью матки и маточных труб. Данная фаза сменяется более длительной второй фазой, в течение которой сперматозоиды со значительно меньшей скоростью транспортируются к месту оплодотворения.
В ампулярной части маточной трубы нормальное количество сперматозоидов в длительной фазе транспорта сохраняется на определенном уровне благодаря непрерывному возмещению уходящих в брюшную полость сперматозоидов за счет тех половых клеток, которые депонированы в нижних отделах полового тракта (крипты шейки матки и др.).
Большое значение в транспорте гамет и оплодотворении имеет так называемый захват яйцеклетки ампулярным отделом маточной трубы. Этот процесс заключается в том, что ампулярный отдел трубы своими фимбриями как бы накрывает яичник в том месте, где произошла овуляция; при этом значительно облегчается перемещение зрелой яйцеклетки в ампулу маточной трубы. Механизм этого феномена окончательно еще не исследован.
Существенным является вопрос о выживаемости сперматозоидов в половых путях женщины. Некоторые авторы полагают, что жизнеспособность спермы сохраняется в течение нескольких (до 5) дней. Однако следует учитывать, что сохранение подвижности сперматозоидов не обязательно свидетельствует об их оплодотворяющей способности.
А В наиболее благоприятных условиях, когда мужские половые клетки находятся в цервикальной слизи на фоне высокого содержания эстрогенов в организме, оплодотворяющая способность сперматозоидов держится до 2 сут после эякуляции во влагалище. В связи с этим принято считать, что для достижения беременности оптимальная частота половых сношений в период до и после овуляции должна быть каждые 2 дня. При более частых половых сношениях фертильность спермы снижается.
Перед оплодотворением зрелая яйцеклетка со всех сторон окружена лучистым венцом (corona radiata). Отчетливо заметна блестящая оболочка (zona pellucida). Уже через несколько минут после разрыва фолликула (овуляции) яйцеклетка попадает в полость маточной трубы. Этому процессу способствует ряд факторов: "захват" ее фимбриями маточной трубы со стороны яичника, в котором произошла овуляция, направление тока фолликулярной жидкости при разрыве фолликула и др. Этим факторам принадлежит очень важная роль в первоначальном транспорте яйцеклетки, которая лишена самостоятельной подвижности. Способность яйцеклетки к оплодотворению в среднем составляет 24 ч. Современные методы диагностики (ультразвуковое исследование, лапароскопия) позволяют не только наблюдать за процессом овуляции, но и фиксировать этот процесс на фотопленку.
Яйцеклетка, попавшая в ампулярный отдел маточной трубы, быстро окружается большим количеством сперматозоидов, являющихся носителями как Х-, так и Y-половых хромосом. Под микроскопом сперматозоиды с Х-хромосомой имеют несколько более крупные размеры, чем несущие Y-хромосому. Сперматозоиды, окружающие яйцеклетку, начинают пенетри-ровать в клетки лучистого венца. Процесс пенетрации обусловлен наличием ряда ферментов, которые содержатся как в головке сперматозоида, так и в трубной жидкости (рис. 3.3).
Сразу же после слияния мембран половых клеток происходит кортикальная реакция яйцеклетки, являющаяся составной частью обеспечения блока полиспермии. После этого хромосомы зиготы вступают в первое митотическое деление, которое наступает через 24 ч после начала оплодотворения. Ядро оплодотворенной яйцеклетки (зиготы) содержит диплоид-

Рис. 3.3. Пенетрация сперматозоидов в яйцеклетку в ампулярном отделе маточной трубы (схема).
ный набор хромосом (46). Таким образом, новый организм является носителем генетической информации обоих родителей.
После оплодотворения (через 24 ч) начинается дробление оплодотворенной яйцеклетки. Первоначально дробление имеет синхронный характер. Через 12 ч от начала возникновения 2 бластомеров возникают 4 бластомера и т.д. К 96 ч от момента слияния ядра сперматозоида с ядром яйцеклетки зародыш состоит из 16-32 бластомеров (стадия морулы). На этой стадии оплодотворенное яйцо (зигота) попадает в матку.
Поскольку дробящаяся яйцеклетка не обладает самостоятельной подвижностью, ее транспорт определяется взаимодействием сократительной активности маточной трубы (основной фактор), движениями цилиарного эпителия эндосальпинкса и капиллярным током жидкости в направлении от ампулярного конца маточной трубы к матке.
Транспорт яйцеклетки по маточной трубе находится под воздействием гормонов. Как известно, после овуляции на месте лопнувшего фолликула образуется новая эндокринная железа - желтое тело. Оно выделяет как прогестерон, так и эстрогенные гормоны. Именно этим двум половым гормонам принадлежит ведущая роль в обеспечении кинетики маточных труб. Под влиянием относительно низкого содержания прогестерона и более высокой концентрации в крови эстрогенов (что имеет место непосредственно после овуляции) повышается тонус ампулярно-перешеечного отдела трубы. В результате яйцеклетка задерживается в ампулярном отделе, где происходит ее оплодотворение и начинается процесс клеточного деления с образованием бластомеров. В дальнейшем происходит постепенное продвижение оплодотворенной яйцеклетки по перешейку маточной трубы к матке. Под влиянием нарастающих концентраций прогестерона желтого тела сократительная функция маточных труб приобретает перистальтический характер, при этом волны сокращений направлены в сторону матки. Происходит расслабление трубно-маточного соединения, и яйцеклетка из маточной трубы попадает в полость матки.
 Эндометрий
Эндометрий
Рис. 3.4. Транспорт оплодотворенной яйцеклетки по маточной трубе вплоть до имплантации (схема).
1 - яйцеклетка в ампуле маточной трубы; 2 - оплодотворение; 3-7 - различные стадии образования бластомеров; 8 - морула; 9, 10 - бластоциста; 11 - имплантация.
Анализируя механизмы гормональной регуляции кинетики маточных труб, следует подчеркнуть, что только определенное соотношение между концентрациями в крови прогестерона и эстрогенов обеспечивает описанное выше четкое физиологическое взаимодействие различных отделов маточной трубы и своевременный транспорт плодного яйца в полость матки. Наряду с прогестероном и эстрогенами в осуществлении физиологической кинетики маточных труб в ранние сроки беременности известное значение отводится таким гормонам, как тестостерон и кортизол. Однако роль этих гормонов изучена в настоящее время еще недостаточно.
Имплантация плодного яйца. Проделав путь по маточной трубе в течение 4 сут, плодное яйцо на стадии морулы попадает в матку, где превращается в бластоцисту (рис. 3.4). Стадия бластоцисты характеризуется тем, что блас-томеры подвергаются определенным изменениям. Часть бластомеров, более крупных по своим размерам, образует так называемый эмбриобласт, из которого в дальнейшем развивается эмбрион. Другая часть клеток, более мелких и располагающихся по периферии плодного яйца, образует питательную оболочку - трофобласт. В дальнейшем наиболее развитая часть трофобласта превращается в плаценту. В полости матки бластоциста приближается к месту имплантации (нидации) (рис. 3.5, а,б). Локализация имплантации имеет свои закономерности и, по-видимому, в значительной степени определяется местными особенностями эндометрия. Обычно бластоциста имплантируется в области передней или задней стенки матки.
Затем начинается погружение бластоцисты в эндометрий, который к этому времени превращается в децидуальную оболочку (рис. 3.5, в,г).
 |
 |
Рис. 3.5. Имплантация.
а - бластоциста перед имплантацией; б - начальный контакт бластоцисты с деци-дуальной оболочкой матки; в - погружение бластоцисты в децидуальную оболочку; г - завершение имплантации.
Децидуальная оболочка представляет собой видоизмененный функциональный слой эндометрия. К моменту имплантации оплодотворенной яйцеклетки слизистая оболочка матки находится в секреторной фазе, железы пилообразно изменены и заполнены секретом, клетки стромы имеют округлый вид и содержат большое количество гликогена, липидов, нейтральных мукополисахаридов, солей и микроэлементов, ферментов и их ингибиторов, иммуноглобулинов и многие другие биологически активные соединения, необходимые для жизнедеятельности зародыша.
Процесс имплантации в первую очередь связан с гормональными факторами. Ведущая роль принадлежит половым (стероидным) гормонам. Во время беременности происходит развитие и активное функционирование желтого тела яичника, возникшего на месте лопнувшего фолликула. Желтое тело секретирует большое количество прогестерона и несколько меньшее - эстрогенов. Эти половые гормоны через специфические стероидные рецепторы, находящиеся в эндометрии, оказывают выраженное воздействие на секреторные преобразования слизистой оболочки матки и процессы ее де-цидуализации. Кроме стероидных, определенную роль в имплантации играют и некоторые другие гормоны (пролактин, глюкокортикоиды).
Децидуальная ткань как своеобразный эндокринный орган имеет прямое отношение к имплантации и дальнейшим стадиям постимплантацион-ного развития зародыша. Установлено, что эндометрий женщины вырабатывает иммунореактивный пролактин. Он начинает секретироваться с 9-го дня после овуляции, при этом во время беременности продукция тканевого пролактина значительно повышается, поэтому в настоящее время некоторые исследователи склонны рассматривать децидуальную ткань как своеобразный эндокринный орган.
Процесс имплантации, который в среднем продолжается около 2 дней, сопровождается не только значительными изменениями клеточных элементов желез и стромы эндометрия, но и выраженными гемодинамическими сдвигами местного характера. Вблизи места имплантации бластоцисты отмечается расширение кровеносных сосудов и образование синусоидов, представляющих собой расширенные капилляры и венулы. Этим процессам принадлежит большая роль в процессах обмена между материнским организмом и зародышем.
Ранний эмбриогенез
После имплантации происходит быстрое развитие как зародыша, так и его оболочек. На трофобласте образуются ворсинки, которые на ранних стадиях онтогенеза равномерно покрывают всю поверхность бластоцисты. Первичные ворсины являются бессосудистыми образованиями. Постепенно наружный слой трофобласта утрачивает клеточные границы, превращаясь в синцитий (симпласт). Внутренний слой трофобласта сохраняет клеточное строение и называется цитотрофобластом. Более подробно дальнейшие процессы дифференциации трофобласта описаны в следующем разделе (см. раздел 3.2).
Одновременно с трофобластом развивается и эмбриобласт. Этот процесс особенно активизируется после имплантации. Клетки эмбриобласта дифференцируются в два узелка: эктобластический и эндобластический. Вскоре в центральных частях этих узелков образуются полости, в результате чего эктобластический узелок превращается в эктобластический пузырек, а эн-
На ранних стадиях развития происходит и дифференциров-ка мезенхимы. С одной стороны клетки мезенхимы оттесняются на периферию - к трофобласту. С другой стороны происходит скопление мезенхимальных клеток вокруг амниотического и желточного пузырьков, а также около зародыша. В результате этого стенки пузырьков становятся двухслойными, а сам зародыш дифференцируется в экто-, эндо- и мезодерму. Эти три зародышевых листка служат исходным материалом для формирования в дальнейшем всех органов и тканей эмбриона и плода.
По мере дальнейшего внутриутробного развития происходит быстрое увеличение размеров амниотического пузырька, который превращается в амниотическую полость. В полости наблюдается быстрое накопление прозрачной жидкости (амниотическая жидкость), при этом стенка амниотического пузырька приближается к ворсинчатой оболочке и, наконец, сливается с ней. Зародыш начинает вворачиваться в полость амниона, а желточный пузырек атрофируется.
Одновременно с развитием плодных оболочек из каудального отдела первичной кишки зародыша образуется выпячивание - аллантоис. По ал-лантоису сосуды зародыша "подрастают" к ворсинчатой оболочке, врастая затем в каждую ворсину. В результате этого бессосудистый хорион васкуля-ризируется.
После завершения начальных этапов онтогенеза эмбрион окружен ам-ниотической жидкостью и тремя оболочками: децидуальной, ворсинчатой и водной.
Децидуальная оболочка представляет собой трансформированный в связи с беременностью функциональный слой эндометрия. К моменту имплантации эндометрий находится в секреторной фазе. Он состоит из двух слоев: компактного и спонгиозного. Компактный слой, обращенный в полость матки, содержит выводные протоки маточных желез и клетки стромы эндометрия, превратившиеся во время беременности в децидуальные. Спон-гиозный слой в основном состоит из желез.
В процессе роста плодного яйца decidua parietalis и decidua capsularis растягиваются, истончаются и приближаются друг к другу. На 4-5-м месяце беременности плодное яйцо занимает уже всю полость матки, и decidua parietalis сливается с decidua capsularis. Наоборот, decidua basalis значительно гипертрофируется и превращается в материнскую часть плаценты. В этой части плаценты находятся многочисленные ворсины хориона, вокруг которых формируется межворсинчатое пространство.
Децидуальная оболочка является для плода питательным и защитным слоем: трофическая функция ее в основном осуществляется на ранних этапах внутриутробного развития, защитная роль наиболее полно проявляется высокой фагоцитарной активностью. Децидуальная ткань лизирует
микроорганизмы и инактивирует их токсины, принимает также участие в синтезе углеводов, липидов и белков. В ней происходит синтез пролактина и простагландинов. Таким образом, децидуальной оболочке принадлежит очень важная роль в имплантации и дальнейшем развитии эмбриона и плода.
Амнион (водная оболочка) обращена к плоду. Она выстилает плаценту и переходит на пуповину, сливаясь в области пупочного кольца с кожей плода. Макроскопически амнион представляет собой тонкую полупрозрачную мембрану. В процессе эмбриогенеза амнион развивается из эктобластического пузырька. Из эктодермы формируется эпителий амниона, из мезодермы - соединительнотканная основа.
На ранних стадиях развития эпителий амниона представлен крупными полигональными клетками, с 3-го месяца беременности он становится кубическим. Эпителиальные клетки амниона содержат липиды, полисахариды,
протеины, фосфорные соединения, а также ряд ферментов, участвующих в процессах метаболизма и обмене стероидных гормонов.
Амнион вместе с гладким хорионом принимает активное участие в обмене околоплодных вод, а также в параплацентарном обмене. По своим физическим свойствам плодные оболочки отличаются друг от друга. Так как амниотическая оболочка очень плотная и выдерживает давление в несколько раз большее, чем гладкий хорион, в родах разрыв гладкого хориона наступает раньше, чем амниона.
ПЛАЦЕНТА
Плацента человека имеет гемохориальный тип строения. Этот тип плацен-тации характеризуется наличием непосредственного контакта материнской крови с хорионом вследствие нарушения целостности децидуальной оболочки матки со вскрытием ее сосудов.
Возникновение гемохориальной плаценты в эволюции является высшей стадией. В ней отражены сложнейшие взаимоотношения функциональных систем матери и плода.
Развитие и функции плаценты
Основной частью плаценты являются ворсины хориона - производные тро-фобласта. На ранних этапах онтогенеза трофобласт образует протоплазма-тические выросты, состоящие из клеток цитотрофобласта (первичные ворсины). Первичные ворсины не имеют сосудов, и поступление питательных веществ и кислорода к организму зародыша из окружающей их материнской крови происходит по законам относительно простых законов осмоса и диффузии. К концу 2-й недели беременности в первичные ворсины врастает соединительная ткань и образуются вторичные ворсины. Их основу составляет соединительная ткань, а наружный покров представлен эпителием (трофобласт). Как первичные, так и вторичные ворсины равномерно распределяются по поверхности плодного яйца.
Эпителий вторичных ворсин состоит из двух слоев: слоя цитотрофобласта (слой Лангханса) и синцития (симпласта). Слой цитотрофобласта состоит из клеток округлой формы со светлой цитоплазмой. Ядра клеток крупные. В синцитии границы клеток практически неразличимы, цитоплазма темная, зернистая, с щеточной каймой. Ядра относительно небольших размеров, шаровидной или овальной формы.
С 3-й недели развития зародыша начинается очень важный процесс развития плаценты, который заключается в васкуляризации ворсин и превращении их в третичные, содержащие сосуды. Формирование сосудов плаценты происходит как из ангиобластов зародыша, так и из пупочных сосудов, растущих из аллантоиса.
Сосуды аллантоиса врастают во вторичные ворсины, в результате чего каждая вторичная ворсина получает васкуляризацию. Так осуществляется важнейший процесс внутриутробного развития - васкуляризация хориона. Установление аллантоидного кровообращения обеспечивает интенсивный обмен между организмами плода и матери.
▲ Превращение вторичных ворсин в третичные рассматривается как важнейший критический период эмбрионального развития, так как аллантоис обладает очень высокой чувствительностью к действию повреждающих факторов окружающей среды. Его повреждение сопровождается гибелью сосудов, в результате чего прекращается важнейший процесс - васкуляризация хориона, а это в свою очередь приводит к гибели зародыша на самых ранних сроках.
На ранних стадиях внутриутробного развития ворсины хориона равномерно покрывают всю поверхность плодного яйца. Однако начиная со 2-го месяца онтогенеза на большей поверхности плодного яйца ворсины атрофируются, в то же время пышно развиваются ворсины, обращенные к базальной части децидуальной оболочки. Так формируются гладкий и ветвистый хорион.
Дальнейшее развитие и дифференцировка хориона характеризуются следующими основными моментами. При сроке беременности 5-6 нед толщина синцитиотрофобласта превосходит толщину слоя Лангханса, а, начиная со срока 9-10 нед синцитиотрофобласт постепенно истончается и количество ядер в нем увеличивается. На свободной поверхности синцитиотрофобласта, обращенной к межворсинчатому пространству, становятся хорошо заметными длинные тонкие цитоплазматические выросты (микроворсины), которые значительно увеличивают резорбционную поверхность плаценты. В начале II триместра беременности происходит интенсивное превращение цитотрофобласта в синцитий, в результате чего на многих участках слой Лангханса полностью исчезает.
В конце беременности в плаценте начинаются инволюционно-дистрофические процессы, которые иногда называют старением плаценты. Из крови, циркулирующей в межворсинчатом пространстве, начинает выпадать фибрин (фибриноид), который откладывается преимущественно на поверхности ворсин. Выпадение этого вещества способствует процессам микро-тромбообразования и гибели отдельных участков эпителиального покрова ворсин. Ворсины, покрытые фибриноидом, в значительной степени выключаются из активного обмена между организмами матери и плода.
Происходит выраженное истончение плацентарной мембраны с 25 мкм в начале беременности до 5 мкм в конце ее. Строма ворсин становится более волокнистой и гомогенной. Наблюдается некоторое утолщение эндотелия капилляров. В участках дистрофии нередко откладываются соли извести. Все эти изменения отражаются на основных функциях плаценты (дыхательной, трофической, обменной, эндокринной и др.).
Наряду с процессами инволюции в плаценте на протяжении беременности наблюдаются и явления противоположного свойства. Происходит увеличение молодых ворсин, которые в значительной мере компенсируют функцию утраченных. Однако развитие молодых ворсин лишь частично улучшает функцию плаценты в целом. В результате этого в конце беременности наблюдается снижение функции плаценты.
Строение зрелой плаценты (рис. 3.8). Макроскопически зрелая плацента очень напоминает толстую мягкую лепешку. Масса плаценты составляет 500-600 г, диаметр 15-18 см, толщина 2-3 см. Плацента имеет две поверхности: материнскую, обращенную к стенке матки, и плодовую - в сторону плода.

Рис. 3.8. Внешний вид зрелой плаценты, а - материнская часть; б - плодовая часть.
Материнская поверхность плаценты имеет серовато-красный цвет и представляет собой остатки базальной части децидуальной оболочки.
Плодовая поверхность сверху покрыта блестящей амниотической оболочкой, под которой к хориону подходят сосуды, идущие от места прикрепления пуповины к периферии плаценты. Основная часть плодовой плаценты представлена многочисленными ворсинами хориона, которые объединяются в дольчатые образования - котиледоны, или дольки. Их число достигает 15-20. Дольки плаценты образуются в результате разделения ворсин хориона перегородками (септами), исходящими из базальной пластинки. К каждой из таких долек подходит свой крупный сосуд.
Микроскопическое строение зрелой ворсины. Принято различать два вида ворсин: свободные и закрепляющие (якорные). Свободные ворсины, а таких большинство, погружены в межворсинчатое пространство децидуальной оболочки и "плавают" в материнской крови. В противоположность им якорные ворсины прикреплены к базальной децидуальной оболочке и обеспечивают фиксацию плаценты к стенке матки. В третьем периоде родов связь таких ворсин с децидуальной оболочкой нарушается и под влиянием маточных сокращений плацента отделяется от стенки матки.
При микроскопическом изучении строения зрелой ворсины удается дифференцировать следующие образования (рис. 3.9):
Синцитий, не имеющий четких клеточных границ;
Слой (или остатки) цитотрофобласта;
Строму ворсины;
Эндотелий капилляра, в просвете которого хорошо заметны элементы
крови плода.
При электронной микроскопии ворсин хориона было установлено, что синцитий имеет на своей поверхности многочисленные микроворсины, которые значительно увеличивают обменную поверхность плаценты.
Маточно-плацентарное кровообращение. При наличии пла-
Кровоток в матке осуществляется с помощью 150-200 материнских спиральных артерий, которые открываются в обширное межворсинчатое пространство. Спиральные артерии имеют своеобразное строение, их стенки лишены мышечного слоя, а устья не способны сокращаться и расширяться.
Спиральные артерии обладают низким сосудистым сопротивлением току крови. В противоположность маточным артериям, в которых выраженное снижение сосудистого сопротивления наблюдается с 12-13 нед беременности, в спиральных артериях, как это было установлено с помощью допплерометрии, этот процесс имеет место уже с 6 нед беременности. Наиболее выраженное снижение сосудистого сопротивления в спиральных артериях наблюдается в 13-14 нед беременности, что морфологически отражает завершение процесса инвазии ворсин трофоблас-та в децидуальную оболочку.
Описанные особенности гемодинамики имеют очень большое значение в осуществлении бесперебойного транспорта артериальной крови от организма матери к плоду. Излившаяся артериальная кровь омывает ворсины хориона, отдавая при этом в кровь плода кислород, необходимые питательные вещества, многие гормоны, витамины, электролиты и другие химические вещества, а также микроэлементы, необходимые плоду для его правильного роста и развития. Кровь, содержащая СО 2 и другие продукты метаболизма плода, изливается в венозные отверстия материнских вен, общее число которых превышает 180.
Кровоток в межворсинчатом пространстве в конце беременности достаточно интенсивен и в среднем составляет 500-700 мл крови в минуту.
Особенности кровообращения в системе мать -плацента -плод. Артериальные сосуды плаценты после отхождения от пуповины делятся радиально в соответствии с числом долек плаценты (котиледонов). В результате дальнейшего разветвления артериальных сосудов в конечных ворсинах образуется сеть капилляров, кровь из которых собирается в венозную систему. Вены, в которых течет артериальная кровь, собираются в более крупные венозные стволы и наконец впадают в вену пуповины (рис. 3.10).
 |
Рис. 3.10. Кровообращение в системе мать - плацента - плод.
1 - миометрий; 2 - свободные ворсины; 3 - якорная ворсина; 4 - децидуальная оболочка; 5 - межворсинчатое пространство; 6 - спиральные артерии; 7 - хориаль-ная пластинка; 8 - хориальный эпителий; 9 - амниотический эпителий; 10 -пуповина; 11 - вена пуповины; 12 - артерии пуповины; 13 - отложение фибриноида.
Кровообращение в плаценте поддерживается сердечными сокращениями матери и плода. Важная роль в стабильности этого кровообращения также принадлежит механизмам саморегуляции маточно-плацентарного кровообращения.
Основные функции плаценты. Плацен
ЭМБРИОЛОГИЯ.
ЭМБРИОЛОГИЯ №1
Понятие прогенеза и эмбриогенеза. Периоды и основные стадии эмбриогенеза у человека. Половые клетки человека, их структурно- генетическая характеристика.
ЭМБРИОГЕНЕЗ –
ПРОГЕНЕЗ – период развития и созревания половых клеток – яйцеклеток и сперматозоидов, в результате в зрелых половых клетках возникает гаплоидный набор хромосом, формируются структуры, обеспечивающие их способность к оплодотворению и развитию нового организма.
ПОЛОВЫЕ КЛЕТКИ – ПРИЗНАКИ:
Гаплоидный набор хромосом
Измененная ядерно-цитоплазматическое отношение – отношение объема ядра и цитоплазма
Изменен метаболизм клетки
Клетки высоко дифференцированы (не способны делиться)
ЯЙЦЕКЛЕТКА (открыта Бером) – имеет оолемму, ооплазму (цитоплазму), ядро; органоиды развиты все за исключением центриолей, из включений преобладает желток.
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ: ядро неактивно ни в отношении транскрипции, ни в отношении репликации, т.е. ведет себя пассивно; яйцеклетка накапливает ферменты, факторы и гликоген.
СТРУКТУРНЫЕ ОСОБЕННОСТИ: имеет кортикальный слой цитоплазмы – периферическая гиалоплазма с кортикальными гранулами (мукополисахариды, белки, ферменты); полярна – выделяют два полюса: анимальный (сосредоточены органоиды) и вегитативный (содержит < или >количество белка)
У человека маложелтковая, вторичноолиголецитальная, изолецитальная
Яйцеклетку окружают прозрачная (лецитальная) оболочка – zona pellucida (ее образуют Zp белки). Среди них есть Zp2 белок – препятствует полиспермии, Zp3 рецептор к сперматозоиду.
Яйцеклетка окружена фолликулярными клетками, которые доставляют к ней питание – формируют лучистый венец.
СПЕРМАТОЗОИД – выделяют четыре отдела: головка (содержит крупное ядро и акросому – видоизмененная лизосома), шейка(проксимальная центриоль), тело (митохондриальные спирали и дистальная центриоль), хвостик (представлен жгутиком)
Максимальная способность к оплодотворению до двух суток.
Направленная миграция сперматозоидов определяется хемотаксисом и реотаксисом, важными показателями при этом являются рН и слизь. Происходит капоцитация – под действием секретов женских половых путей сперматозоид приобретают оплодотворяющие способности.
Продвижение сперматозоида облегчают простогландины (действуют на оболочку маточных труб)
ЭМРИОЛОГИЯ №2
Основные стадии эмбриогенеза. Понятие оплодотворения. Характеристика оплодотворения у человека: морфология, необходимые условия. Понятие зиготы.
ЭМБРИОГЕНЕЗ – эмбриональное развитие человека. Продолжается 280 дней, делится на три периода: начальный (первая неделя развития), зародышевый (2-8 неделя развития – закладка основных органов), плодный (9неделя – до рождения).
Ранний эмбриогенез делится на стадии:
ЗИГОТА – начало синтеза ДНК и белка
ДРОБЛЕНИЕ – начало синтеза основных типов РНК
МОРУЛА – клетки зародыша тотипотентны (взаимозаменяемы)
БЛАСТОЦИСТА – происходит утрата тотипотентности и клетки детерминируются к образованию зародышевых и внезародышевых структур.
ГАСТРУЛА – появляются зародышевые листки и стволовые клетки
ОРГАНОГЕНЕЗ – из ткани формируются органы, идет формирование зачатков органов из клеточных клонов
ОПЛОДОТВОРЕНИЕ – слияние мужской и женской половых клеток, в результате чего восстанавливается диплоидный набор хромосом, характерный для данного вида и возникает качественно новая клетка – зигота (оплодотворенная яйцеклетка или одноклеточный зародыш)
Три стадии:
ПОЛОЖИТЕЛЬНЫЙ РЕОТАКСИС – движение сперматозоида против тока жидкости, выделяемой маточными трубами
ХЕМОТКСИС – половые клетки выделяют гормоны (гамоны); ♀ - гиногамоны, у ♂ - андрогамоны.
КАПОЦИТАЦИЯ – приобретение сперматозоидом оплодотворяющей способности под действием секрета маточных труб
АКТИВАЦИЯ ЯЙЦЕКЛЕТКИ – участок мембраны яйцеклетки, полученный от сперматозоида проницаем для ионов Na→изменяется потенциа клетки→ионы Ca выходят из клеточного депо в цитоплазму→экзоцитоз кортикальных гранул. Это приводит к образованию оболочки оплодотворения – изменение свойств блестящей оболочки, она препятствует полиспермии.
СПЕРМАТОЗОИД ВНУТРИ ЯЙЦА – длится 12 часов, в результате образуется зигота. В этот момент ядра половых клеток называют пронуклеус. Их ядерный матрикс разрыхляется, оболочки исчезают – стадия синкариона. Пронуклеосы сближаются, в каждом из них происходит удвоение ДНК и образование хромосом, которые перемешиваются и выстраиваются в метафазную пластинку первого деления мейоза.
ДИСТАНТНОЕ ВЗАИМОДЕЙСТВИЕ
КОНТАКТНОЕ ВЗАИМОДЕЙСТВИЕ – сперматозоид раздвигает лучистый венец и образует канал в блестящей оболочке. Сперматозоид контактирует с рецептором (Zp3 белок) и начинается акросомная реакция – экзоцитоз содержимого акросомы для локального разрушения прозрачной оболочки. Внутри головки происходит: внутрь ионы Ca и Na, наружу H2. Результат – увеличение концентрации Ca запускает ряд процессов, ведущих к увеличению внутриклеточного pH, а это запускает акросомную реакцию. Результат акросомной реакции – образование канала в прозрачной оболочке, через который проходит сперматозоид. В оолемму встраивается мембрана сперматозоида.
ТРЕТЬЯ СТАДИЯ
ЗНАЧЕНИЕ СПЕРМАТОЗОИДА – ½ хромосом в зиготе отцовские; митохондриальный геном отца; вносит сигнальный белок дробления; снимается блок мейоза; определяется генетический по организма.
Следствием оплодотворения являются изменение объема зиготы, деполяризация плазматической мембраны и образование оболочки оплодотворения.
ЭМБРИОЛОГИЯ №3
Понятие дробления зародыша. Характеристика дробления человека: типы дробления, время эмбриогенеза, продолжительность, условия. Строение зародыша на стадии имплантации у человека.
ДРОБЛЕНИЕ – последовательное митотическое деление зиготы на клетки (бластомеры) без роста дочерних клеток до размера материнской.
Разделившиеся клетки не растут, не расходятся, сохраняют диплоидность. Сокращается х митотический цикл за счет G1 и G2 периодов.
Суть дробления : образуется многоклеточный организм, восстанавливаются ядерно-цитоплазматические отношения.
У человека дробление: полное (материал зиготы дробится полностью), асинхронное (увеличение числа бластомеров происходит с нарушением геометрической прогрессии.
ВИДЫ БЛАСТОМЕРОВ: мелкие светлые и крупные темные
Через 30 часов проходит первая борозда дробления (образуются два бластомера)
Через 40 часов – 4 бластомера
Через 50-60 – образуется морула (тутовая ягода)
Мелкие светлые – по периферии, образуют трфобласт
Крупные темные – внутри, образуют эмбриобласт.
На 5 сутки образуется бластула (бластоциста). Ее особенность – бластоцель.
В начале из эмбриобласта образуется зародышевый узелок, а затем зародышевый щиток.
На 5 сутки бластоциста попадает в матку.
На 6-7 сутки протекает первая фаза гаструляции и происходит имплантация.
ИМПЛАНТАЦИЯ – внедрение зародыша в слизистую оболочку матки, две стадии:
ПРИЛИПАНИЕ (адгезия) – клетки трофобласты начинают активно делиться и сливаясь образуют симпластотрофобласт – ворсину хориона.
ПРОНИКНОВЕНИЕ (инвазия) – симпластотрофобласт выделяет протолитические ферменты, которые последовательно разрушают эпителий, соединительную ткань и кровеносные сосуды слизистой оболочки матки.
Первоначально зародыш питается разрушенными тканями матери – гистотрофный тип питания, а затем питается материнской кровью – гематрофный тип питания.
ЭМБРИОЛОГИЯ №4
Понятие и основные механизмы гаструляции. Типы гаструляции. Морфологическая и временная характеристика гаструляции у человека. Строение двухнедельного зародыша человека. Представления о критических периодах развития.
ГАСТРУЛЯЦИЯ – сложный процесс химических и морфогенетических изменений, сопровождающийся размножением, ростом, направленным перемещением и дифференцировкой клеток, в результате образуются зародышевые листки: наружный – эктодерма, внутренний – энтодерма, средний – мезодерма. Это источники зачатков тканей и органов, комплекса осевых органов.
СПОСОБЫ ГАСТРУЛЯЦИИ:
ИНВАГИНАЦИЯ (впячивание) – дно подтягивается к крыше
ЭПИБОЛИЯ (обрастание) – характерна для крупных клеток, переполненных желтком.
ИММИГРАЦИЯ (выселение)
ДЕЛЯМИНАЦИЯ (расщепление)
У человека протекает в две фазы:
ДЕЛЯМИНАЦИЯ – зародышевый щиток (эмбриобласт) расщепляется на эпибласт (все зачатки кроме энтодермы) и гипобласт (энтодерма).
ИММИГРАЦИЯ (на 14-17 сутки) – движение клеточных масс от головного и хвостового концов, затем они идут параллельно, образуя первичную полоску. Материал, который дремал в головном конце приходит в движение и идет навстречу первому потоку. Клеточный материал перестраивается и образуется первичный узелок с ямкой в центре. Через первичный (Гензеновский) узелок перемещается прехордальный зачаток и уходит в энтодерму (в головной конец зародыша); хордальный зачаток через первичный узелок ложится между экто- и энтодермой, образуя хорду. Мезодерма двумя крыльями подворачивается через края первичной полоски и уходит внутрь, располагаясь по бокам от хорды. Эктодерма всегда перемещается пластом, мезодерма – совершает амебообразные движения, энтодерма относительно неподвижна.
ФАКТОРЫ ВЫЗЫВАЮЩИЕ ГАСТРУЛЯЦИЮ:
Неравномерный рост в различных областях зародыша
Механический (бластомеры легко изменяют форму)
Разная интенсивность поглощения воды
Различная пластичность клеточных мембран
Способность клеток к фагоцитозу
Межклеточные взаимодействия
ЭМБРИОЛОГИЯ №5.
Понятие дифференцировки зародышевых листков. Представление об индукции как факторе, вызывающем дифференцировку. Дифференцировка зародышевых листков и образование зачатков тканей и органов у зародыша человека.
ДИФФЕРЕНЦИРОВКА ПЕРВИЧНОЙ ЭКТОДЕРМЫ:
Нервная трубка (нейроциты и нейроглия сетчатки глаза и органа обоняния, нейроциты и нейроглия головного и спинного мозга)
Нервный гребень и ганглиозные пластинки (нейроциты и нейроглия спинальных и вегетативных ганглиев, хромаффинная ткань и мозговое в-во надпочечников)
Плакоды (эпителиальные элементы внутреннего уха)
Кожная эктодерма (эпидермис и его производные, эпителий роговицы глаза, эпителий органов ротовой полости и его производные, эпителий анального отдела прямой кишки, эмаль и кутикула зуба, эпителиальная выстилка влагалища)
Прехордальная пластинка (эпителий ротовой полости и пищевода, эпителий трахеи, бронхов и легких)
Зародышевая эктодерма
Внезародышевая эктодерма (эпителий амниона и пупочного канатика)
ДИФФЕРЕНЦИРОВКА ПЕРВИЧНОЙ ЭНТОДЕРМЫ
Энтодерма кишечной трубки (эпителий кишечника, желудка, печени,поджелудочной)
Желточная энтодерма (эпителий аллантоиса и желточного места)
Зародышевая энтодерма
Внезародышевая энтодерма
ДИФФЕРЕНЦИРОВКА МЕЗОДЕРМЫ
Сомиты
Миотом (скелетная мышечная ткань)
Склеротом (хрящевая и костная ткань)
Дерматом (соединительнотканная основа кожи)
Мезенхима (экзоцелломический эпителий, соединительная ткань желточного мешка амниона и хориона)
Спланхнотом
Париетальный и висцеральный листки
Зародышевая мезодерма
Сегментная ножка нефрогонотома (эпителий гонад и семявыносящих путей и почек)
Парамезонефральный проток (эпителиальная выстилка влагалища, матки, яйцеводов)
Спланхнотом (поперечнополосатая мышечная ткань сердца, корковое в-во надпочечников, мезотелий)
Мезенхима (микроглия, гладкая мышечная ткань, сосуды, соединительные ткани, к-ки крови и кроветворных органов)
Внезародышевая мезодерма
Индукторы – возникают в определенных участках зародыша, обуславливают развитие других участков в определенном направлении.
ЭМБРИОЛОГИЯ №6.
Понятие и значение внезародышевых органов. Их появление в эволюции. Внезародышевые органы у человека. Образование, строение, значение амниона, желточного мешка, аллантоиса. Туловищная складка, ее образование, роль.
ВНЕЗАРОДЫШЕВЫЕ (провизорные, временные) ОРГАНЫ – развиваются в процессе эмбриогенеза вне тела зародыша, выполняют разнообразные функции, обеспечивающие рост и развитие самого зародыша.
Некоторые из этих органов, окружающих зародыш, называют зародышевыми оболочками. К этим органам относятся: амнион, желточный мешок, аллантоис, хорион, плацента
У человека эти органы закладываются рано. К 11 суткам развития начинается выселение мезодермы, которая заполняет полость бластоцисты. Из эпибласта выселяется внезародышевая мезодерма, которая прорастает перед эпибластом и образует закладку будущего амниотического пузырька. Затем по ней прорастает внезародышевая эктодерма. Из эпибласта выселяется мезенхима, которая прорастает перед гипобластом и образует закладку будущего желточного мешка. Позже по ней прорастает внезародышевая энтодерма и образуется желточный мешок.
АЛЛАНТОИС – развивается на 15 сутки эмбриогенеза как выпячивание стенки кишечной трубки. Проводит кровеносные сосуды к ворсинам хориона и, редуцируясь, войдет в состав пупочного канатика.
ХОРИОН – к концу 2 недели эмбриогенеза трофобласт начинает образовывать первичные ворсины хориона. В начале 3 недели к ним подрастает мезенхима и образуются вторичные ворсины хориона. Вскоре мезенхима дифференцируется в соединительную ткань и кровеносные сосуды, так формируются вторичные ворсины хориона, которые сформируют плодную часть плаценты. Ворсины хориона, прилежащие к основной отпадающей оболочке, интенсивно разрастаются и образуется ветвистый хорион, который соединяясь с основной отпадающей оболочкой образует плаценту.
ЖЕЛТОЧНЫЙ МЕШОК – принимает участие в кроветворении до 7-8 недели развития. Здесь образуются первичные половые клетки, позже идет его эволюция и он обнаруживается в составе пупочного канатика в виде узкой трубочки.
АМНИОН – достигает большого развития, создает благоприятную водную среду для развития зародыша
???????????????????????????????????????????????????????????????????????????????????????????????????
ЭМБРИОЛОГИЯ №7
Плацента. Ее значение, появление в эволюции. Типы плацент. Плацента человека: тип, строение, функции. Структура и значение плацентарного барьера.
ПЛАЦЕНТА – ворсины хориона прилежащие к основной отпадающей оболочке интенсивно разрастаются и образуется ветвистый хорион, который соединяется с основной отпадающей оболочкой образуя плаценту или детское место.
Зрелая плацента в диаметре 17-20 см. Толщина 2-4 см. Вес 500-600 г.
По строению дискоидальная, гемохориальная, ворсинковая.
ТИПЫ ПЛАЦЕНТ:
В связи с особенностями проникновения ворсин хориона в матку делят:
эпителиальнохориальная – ворсины хориона не разрушают эпителий матки (лошади, свиньи)
десмохориальная – ворсины хориона разрушают эпителий матки и контактируют с соединительной тканью (жвачные и парнокопытные)
эндотелиохориальная – ворсины хориона разрушают эпителий матки, соединительную ткань и контактируют с эндотелием сосудов (хищники и ластонгие)
гемохориальная – ворсины разрушают эпителий матки, соединительную ткань и стенки сосудов и непосредственно контактируют с материнской кровью (приматы, человек)
По характеру питания:
Хорион поглощает из материнского организма белки, расщепляет их до аминокислот. Синтез эмбриоспецифических белков происходит в печени эмбриона. Сюда относятся эпителиохориальные и десмохориальные плаценты. Детеныши после рождения способны к самостоятельному питанию и передвижению.
Хорион поглощает аминокислоты, синтезирует эмбриоспецифические белки (все остальные плаценты) детеныши после рождения сравнительно долго адаптируются.
СТРОЕНИЕ ПЛАЦЕНТЫ:
Плодная часть
АМНИОТИЧЕСКАЯ ОБОЛОЧКА – состоит из однослойного призматического эпителия и внезародышевой соединительной ткани.
ХОРИАЛЬНАЯ ПЛАСТИНКА
ХОРИАЛЬНЫЕ ВОРСИНЫ
Строму (основу) хориальной пластины и ворсин составляет рыхлая соединительная ткань (содержит аморфное вещество, аргирофтльные волокна, множество капилляров и клетки Кащенко-Ховбауэра – примитивные макрофаги). Сверху пластины покрыты трофобластным эпителием. В первом триместре покрыты цитотрофобластом, во втором покрыты цито- и симпластотрофобластом. В третьем – фибриноидом (гомогенная оксифильная масса, которая является продуктом сыворотки крови и распада трофобластных элементов). В ходе беременности ворсины сильно увеличиваются в размерах и возрастает их ветвление. Они собираются группами по 15-16 штук и формируют котиледоны (группа ворсин, связанная с помощью ствола ворсин с хориальной пластинкой). Выделяют якорные ворсины (соприкасаются с материнской тканью) и конечные ворсины (соприкасаются с материнской кровью). Образуется соединительная ткань: септы на границе между котиледонами.
Материнская часть
ОСНОВНАЯ ОТПАДАЮЩАЯ ОБОЛОЧКА (базальная пластина)
СОЕДИНИТЕЛЬНОТКАННЫЕ СЕПТЫ
ЛАКУНЫ, ЗАПОЛНЕННЫЕ МАТЕРИНСКОЙ КРОВЬЮ
В соединительнотканной слизистой оболочке появляются децидуальные клетки (те же соединительнотканные клетки крупных размеров и богатые гликогеном. Увеличивается просвет сосудов и гидратация слизистой, появляется исключительная фагоцитарная активность
ФУНКЦИИ ПЛАЦЕНТЫ:
Синтез БЖУ
Ионы Ca, Fe, Zn, Mg, Cu, P
Депонирует и содержит витамины групп: A, B1, B2, B6, C, D, E
Факторы свертывания крови
Дыхательная
Трофическая
Эндокринная – все гормоны синтезируются симпластотрофобластом. Они необходимы для роста и развития плода, готовят организм матери к родам и лактации.
Гормоны :
хорионический гонадотропин (поддержание желтого тела, выделяется с первых дней беременности, действует на желтое тело и стимулирует секрецию прогестеронов и эстрогенов, которые действуют на эндометрий, вызывая рост зародыша
хорионический соматомаммотропин (действует на молочную железу и желтое тело, вызывая изменения в организме матери)
кортиколиберин (определяет срок наступления родов)
Иммунологическая защита
МЕХАНИЗМЫ ЗАЩИТЫ ПЛОДА: симпластотрофобласт – синтезирует белки тормозящие иммунный ответ матери, гормоны плаценты – угнетают материнские лимфоциты, фибриноид плаценты
Барьерная (защитная) – выражена слабо, поэтому проникают наркотики, алкоголь, никотин, яды, лекарства (антибиотики, сульфаниламиды, анальгетики), вирусы, бактерии
ПЛАЦЕНТАРНЫЙ БАРЬЕР: барьер между кровью матери и плода. К барьеру подходят: цитотрофобласт с его базальной мембраной, соединительная ткань, строма ворсины, эндотелий кровеносного сосуда и его базальная мембрана. С материнской стороны кровь поступает в межворсинковое пространство через 30 спирально извитых артерий матки по давлением 70-80 мм рт. ст. Материнская кровь омывает ворсины, а затем скапливается на дне плацентарных отсеков, откуда уносится через краевой синус в маточные вены. Объем крови 150 мл.
ЭМБРИОЛОГИЯ №8
Понятие и значение плацентации. Плацентация у человека: временная и морфологическая характеристика. Тип и строение сформированной плаценты.