පෘෂ්ඨවංශික ඉන්ද්රිය පද්ධති. පෘථිවි ඉතිහාසය පෘෂ්ඨවංශී පද්ධතිය
පෘෂ්ඨවංශීන්ගේ මුතුන් මිත්තන්, පෙනෙන විදිහට, තවමත් කර්ණික කුහරයක් අත්පත් කර නොගත් බෙන්ටික්-පෙලාජික් නොවන හිස් කබල නොවන සතුන්ගේ ප්රාථමික ආකාරවලට සමීප විය. අනුමාන වශයෙන්, මේවා කුඩා ජලජ සතුන් වූ අතර, චෝඩේට් වල සාමාන්ය සලකුණු ඇති නමුත් තවමත් ශක්තිමත් අභ්යන්තර ඇටසැකිල්ලක් සහ බලවත් මාංශ පේශි නොතිබුණි. ඔවුන් බොහෝ විට Ordovician හි මුල් කණ්ඩායම් වලින් වෙන් වී ඇත, එනම් අවම වශයෙන් වසර මිලියන 500 කට පෙර. කෙසේ වෙතත්, ඒවා අප නොදන්නා අතර කිසි විටෙකත් සොයා ගැනීමට අපහසුය, මන්ද ඒවායේ කුඩා ප්රමාණය සහ ඝන අස්ථි සැකැස්ම නොමැති වීම නිසා පොසිල සංරක්ෂණය කළ නොහැක.
දුර්වල ලෙස සංරක්ෂණය කර ඇති ප්රාථමික, නමුත්, නිසැකවම, දැනටමත් සම්පුර්ණයෙන්ම පිහිටුවා ඇති පෘෂ්ඨවංශීන් (scutellum agnathans) Ordovician - Lower Silurian තැන්පතු වලින් (වසර මිලියන 450 ක් පමණ පැරණි) දන්නා කරුණකි. ඔවුන් බොහෝ විට මිරිදිය ජලාශවල ජීවත් වූ අතර, ඔවුන්ගේ දේහය ධාරාව මගින් නොගැඹුරු මුහුදු බොක්ක වෙත ගෙන ගොස් තැන්පත් කරන ලදී. ගංගා ඩෙල්ටා සහ මුහුදේ ලවණ ඉවත් කළ ප්රදේශවල ඔවුන්ගේ පැවැත්ම බැහැර නොකෙරේ. පෘෂ්ඨවංශීන්ගේ නටබුන් සාමාන්ය සමුද්ර තැන්පතු වල සිදු වීමට පටන් ගන්නේ ඩෙවෝනියානු මැද සිට පමණි, එනම් වයස අවුරුදු 350 ක පමණ තැන්පතු වල. වසර ගණනාවකට පෙර සහ පසුව. මෙම පාෂාණ විද්යාත්මක දත්ත යෝජනා කරන්නේ පෘෂ්ඨවංශීන් සෑදීම සිදු වූයේ මුහුදේ නොව මිරිදිය ජලයේ බවයි. සමුද්ර chordates - පෘෂ්ඨවංශීන්ගේ මුතුන් මිත්තන් මුහුදේ සිට මිරිදිය දක්වා සංක්රමණය වීමට හේතුව කුමක් විය හැකිද?
රතු ඇස් ඇති ගස් ගෙම්බා(Agalychnis callidryas)
පාෂාණ විද්යාත්මක දත්ත අනුව විනිශ්චය කිරීම, ඕර්ඩොවිසියානු සහ සිලූරියානු යුගයේදී, මුහුදේ සහ සාගරවල පහළ ජෛව සිනෝස් විවිධ සතුන්ගෙන් පොහොසත් විය - පණුවන්, මොලුස්කාවන්, කබොල සහ echinoderms. පතුලේ, බොහෝ විට ටියුනිකේට් ඇතුළු පහත් chordates බොහෝ විය; විශාල cephalopods සහ යෝධ කබොල ගෝනුස්සන් (දිග මීටර් 3-5 දක්වා ළඟා වූ) වැනි බලවත් විලෝපිකයන් ද මෙහි වාසය කළහ. දැඩි එදිරිවාදිකම් ඇති එවැනි සංතෘප්ත ජෛව ජනක තත්වයන් යටතේ නව විශාල කණ්ඩායමක් (පෘෂ්ඨවංශීන්ගේ උප වර්ගයක්) මතුවීමේ හැකියාවක් පෙනෙන්නට නැත. ඒ අතරම, මිරිදිය ජලාශවල දැනටමත් තරමක් පොහොසත් ශාක (ප්රධාන වශයෙන් ඇල්ගී) සහ විවිධ අපෘෂ්ඨවංශීන් සැලකිය යුතු සංඛ්යාවක් පැවතුනි; විශාල සහ ශක්තිමත් විලෝපිකයන් සංඛ්යාවෙන් ස්වල්පයක් විය. එමනිසා, පෘෂ්ඨවංශීන්ගේ මුතුන් මිත්තන් මෙහි විනිවිද යාම සහ ඔවුන්ගේ පසුකාලීන වර්ධනය විය හැකි බව පෙනේ. කෙසේ වෙතත්, මුහුද හා සසඳන විට, සාගර ජීවීන් හඳුන්වාදීම වළක්වන අහිතකර ලක්ෂණ ද මිරිදිය ජලය සතු විය. ජලයේ දියවන ලවණවල නොසැලකිය යුතු ප්රමාණයක් ශරීරයේ අධික ලෙස ජලය දැමීම, ලුණු සංයුතියේ තියුණු උල්ලංඝනයක් සහ පටක වල ඔස්මොටික් පීඩනය හේතුවෙන් සාමාන්ය පරිවෘත්තීය ආබාධයකට තුඩු දිය යුතු ජල පාරගම්ය ආවරණ සහිත සාගර ආක්රමණිකයන්ට තර්ජනයක් විය. මිරිදිය ජලාශ, මුහුදට සාපේක්ෂව, වඩා අස්ථායී රසායන විද්යාව (ඔක්සිජන් අන්තර්ගතයේ තියුණු උච්චාවචනයන් ඇතුළුව) සහ වෙනස් කළ හැකි උෂ්ණත්ව පාලන තන්ත්රය මගින් සංලක්ෂිත වේ. ගංගාවල ජනාවාස සඳහා යම් ප්රදේශයක රැඳී සිටීමට සහ ධාරාව මගින් ප්ලාවනය වීමට ප්රතිරෝධය දැක්වීම සඳහා ඉහළ සංචලතාවක් අවශ්ය විය.
නමුත් විලෝපිකයන්ගේ දුර්වල පීඩනය, ආහාරවල සාපේක්ෂ බහුලත්වය සහ මිරිදිය biocenoses හි අඩු තියුණු තරඟය නිසා chordates හි සමහර නියෝජිතයින් විනිවිද යාමට සහ නව තත්වයන්ට අනුවර්තනය වීමට අවස්ථාවන් නිර්මාණය විය. පෙනෙන විදිහට, පෘෂ්ඨවංශීන් සෑදීම සිදු වූයේ මේ ආකාරයෙන් වන අතර, ඔවුන්ගේ මුතුන් මිත්තන් මුලින්ම මෝයවලට විනිවිද ගොස්, පසුව ගංගාවල පහළ ප්රදේශවලට ගොස් විල් ජනාකීර්ණ කරමින් ඉහළට නැගීමට පටන් ගත්හ. එවැනි ස්ථානවල, ශරීරයෙන් විශාල ජල ප්රමාණයක් පෙරීමට, ශරීරයේ යුෂ වල ඔස්මොටික් පීඩනය උල්ලංඝනය කිරීම සහ නව තත්වයන් තුළ ඌනතාවයෙන් යුත් ලවණ නැතිවීම වැළැක්වීමේ හැකියාව ඇති පෘෂ්ඨවංශීන්ගේ ලක්ෂණ සහිත වකුගඩු මතු විය හැකිය. මිරිදිය ජලාශවල ඔක්සිජන් අන්තර්ගතයේ සෘතුමය උච්චාවචනයන් සඳහා ශ්වසන ඉන්ද්රියයන් වැඩිදියුණු කිරීම අවශ්ය වූ අතර, පෙනෙන විදිහට, ගිලිනු පෙනුම පරිණාමීය ප්රතිචාරයක් විය. ධාරා වල බලය ජය ගැනීමේ අවශ්යතාවය සංචලනය වැඩි දියුණු කිරීමට දායක විය: ශක්තිමත්, නමුත් රඳවා තබා ගැනීමේ නම්යශීලීභාවය, වර්ධනය වූ කොඳු ඇට පෙළේ ස්වරූපයෙන් අක්ෂීය ඇටසැකිල්ලක් - මෝටර් මාංශ පේශිවල වැඩිවන ස්කන්ධය සඳහා ආධාරකයක්. අනෙක් අතට, වැඩි සංචලනය සඳහා ස්නායු පද්ධතිය සහ සංවේදක ඉන්ද්රියයන්, සංසරණ, ආහාර ජීර්ණ සහ බැහැර කිරීමේ පද්ධතිවල සංකූලතා අවශ්ය විය. ශරීරයේ ප්රමාණය වැඩිවීම මගින් මයෝකොර්ඩියල් සංකීර්ණය ශක්තිමත් කිරීම පහසු කරන ලද අතර, එමඟින් චලනය වීමේ වේගය වැඩි කිරීමට හැකි විය, මන්ද දිගු සිරුරක දුර්ලභ කම්පන කෙටි ශරීරයක් ඇති නිරන්තර ඒවාට වඩා යාන්ත්රිකව ප්රයෝජනවත් වේ. ශරීරයේ නිරපේක්ෂ මානයන් වැඩි වීමත් සමඟ ජලයේ චලනය වීමේ වේගය වැඩිවීම මාළු උදාහරණයෙන් පෙන්විය හැකිය.
පළමු පෘෂ්ඨවංශීන්හකු රහිත සම්බන්ධ ( අග්නාතා), පෙනෙන විදිහට නැවුම් මෝයවල සහ විශාල ගංගාවල මුතුන් මිත්තන්ගේ කණ්ඩායම් වලින් වෙන් වී ඇත. මෙය සිදු වූයේ ඕර්ඩොවිසියන් අවසානයේ - සිලුරියන් ආරම්භයේ ය. ඔවුන්ට හකු සහ යුගල කළ වරල් නොතිබුණි, බෙන්තික් ජීවන රටාවකට මඟ පෑදී, ඩෙට්රිටස් සහ සමහර විට, කුඩා, අක්රිය බෙන්ටික් අපෘෂ්ඨවංශීන්ගෙන් පෝෂණය විය. පසුව, මෙම සතුන් ශක්තිමත් පිටත කවචයක් ලබා ගත් අතර, පෙනෙන විදිහට ඔවුන් කොල්ලකාරී ගෝනුස්සන් වැනි මිරිදිය විලෝපිකයන්ගෙන් ආරක්ෂා කරයි - යූරිපෙට්රිඩ් සහ ඒ සමඟම ශරීරයට මිරිදිය විනිවිද යාම අඩු කිරීමට උපකාරී වේ. Ordovician - Silurian හි (ආසන්න වශයෙන් වසර මිලියන 400-450 කට පෙර), අවම වශයෙන් ස්කූටේට් හකු කණ්ඩායම් තුනක් මතු වූ අතර, ඉන් එකකින්, පෙනෙන විදිහට, එකල පෘෂ්ඨවංශීන්ගේ තවත් ශාඛාවක් වෙන් වීමට පටන් ගත්තේය - හකු - Gnathostomata(සන්නද්ධ මාළු - ප්ලැකෝඩර්මි) හකු රහිත මුතුන් මිත්තන්ට වඩා වෙනස්ව, ඔවුන් වේගවත් ගලායාමක් සහිත ගංගාවලට ඇතුළු වී ඇති අතර, තෝරා ගැනීම ශක්තිමත් අභ්යන්තර ඇටසැකිල්ලක් සහ බලවත් මාංශ පේශි සෑදීමට, යුගල වරල් සෑදීමට සහ ඉන්ද්රියයන් තවදුරටත් වැඩිදියුණු කිරීමට දායක විය. සහ මධ්යම ස්නායු පද්ධතිය. මෙය වැඩි සංචලනය සහ වඩා හොඳ දිශානතියක් ලබා දුන් අතර වේගයෙන් ගලා යන ජලයේ විවිධ ආහාර වස්තූන් සඳහා ජීවත් වීමට සහ දඩයම් කිරීමට හැකි විය. ක්රමයෙන්, හකු උපකරණය සාදන ලදී - ජංගම ගොදුර සක්රීයව අල්ලා ගැනීමේ අවයවයකි.
ප්රාථමික පෘෂ්ඨවංශිකයෙකු බව විශ්වාස කෙරේ ( ප්රොටොක්රැනියෝටා) - හකු සහ හකු නැති අයගේ මුතුන් මිත්තන් - හිස් කබලක් මෙන් හිමි ( ඇක්රානියා) සංකීර්ණ සිදුරු සහිත ෆරින්ක්ස් සහිත සහ පෙරහන-පෝෂණය කරන මයික්රොෆේජ් එකක් විය. දැනට ජීවමාන ලම්ප්රේ කීටයන් වන වැලිපණුවන් වැනි ඛණ්ඩනය නොවූ ගිල් ආරුක්කු මගින් ආධාරක වන ෆරින්ක්ස් හරහා ජල ප්රවාහය සපයනු ලැබුවේ ගිල් කුහරය මිරිකන මාංශ පේශි හැකිලීමෙනි (හුස්ම පිටවීම); බිත්ති (චාප) වල ඉලාස්ටික් ලිහිල් කිරීමේදී ආශ්වාස කිරීම සිදු විය; ෆරින්ක්ස් හි ආරම්භක කොටසෙහි ශ්ලේෂ්මල ධාරාවකින් ආහාර අල්ලා ගන්නා ලදී. පරිණාමයේ ඊළඟ අදියරේදී, ගිල් ආරුක්කු ප්රකාශිත ව්යුහයක් ලබා ගත් අතර ඒවාට සම්බන්ධ මාංශ පේශි දිස් වූ අතර, ගිල් කුහරය සක්රීයව පුළුල් කරයි. ඒ සමගම, ජලය ගලා යාමේ වැඩි වීමත් සමඟ විශාල ආහාර අංශු ඉදිරිපස ආරුක්කු මගින් රඳවා තබා ගත හැකිය. අවසාන වශයෙන්, හකු පෘෂ්ඨවංශීන් සෑදීමේදී, ඉදිරිපස ගිල් ආරුක්කු වලින් එකක පහළ (මැන්ඩිබුලර්) කොටස (සමහර විට තුන්වන) පහළ හකු බවට පරිවර්තනය වූ අතර, මෙම ආරුක්කුවේ ඉහළ කොටස (පලටයින්-චතුරස්රය) සවි කර ඇත. අක්ෂීය හිස් කබල, ග්රහණය කර ගැනීමේ අවයවයක් සාදයි - හකු. ඉදිරිපස ආරුක්කු තොල් කාටිලේජ බවට පත්ව ඇත (කාටිලේජිනස් මාළු එය ඇත), පසුපස ආරුක්කු ගිල් උපකරණ සඳහා ආධාරකයක් ලෙස සංරක්ෂණය කර ඇත ( ලෙසර්ටිසර්, රොබින්, 1970).
දැනටමත් මැද ඩෙවෝනියානු (එනම්, වසර මිලියන 320 කට පමණ පෙර), උභයජීවීන් ( උභය ජීවියා) කාබොනිෆරස් යුගයේදී, ඔවුන් විවිධ ප්රමාණවලින්, ව්යුහයෙන් සහ පෙනුමෙන් යුත් කණ්ඩායම් කිහිපයක් විසින් නියෝජනය කරන ලද අතර, ඔවුන් මිරිදිය ජලාශවල වෙරළබඩ කලාපවල පදිංචි වූ නමුත් පසුව ඒවායින් බොහොමයක් අතුරුදහන් විය; ට්රයැසික් යුගයේදී (වසර මිලියන 170-180 කට පමණ පෙර), විශාල උභයජීවීන් - ස්ටෙගොසෙෆල් - මිය ගියේය.

කාබොනිෆරස් යුගයේ මැද (වසර මිලියන 250-260 කට පමණ පෙර), උරගයින් උභයජීවීන්ගෙන් වෙන් විය ( උරගයා) සමස්ත මෙසෝසොයික් යුගයේදී, වසර මිලියන 120 කට වැඩි කාලයක් ඔවුන් පෘථිවියේ ආධිපත්යය දැරූ අතර, නැවුම් සහ සාගර ජලය සහ වාතය ඇතුළුව ජීවිතයේ සෑම අංශයක්ම පාහේ සාර්ථකව ප්රගුණ කළහ. විවිධ වාසස්ථානවල ජීවත් වීමට අනුවර්තනය වූ උරගයන් උප කාණ්ඩ 6-7 කට බෙදා ඇත. උරගයින්ගේ මල් පිපීම පුරාණ උභයජීවීන්ගේ වඳවීම කලින් තීරණය කළේය. ක්රිටේසියස් යුගය අවසන් වන විට (වසර මිලියන 60-80 කට පමණ පෙර), බොහෝ උරග කණ්ඩායම් ද සාපේක්ෂව ඉක්මනින් මිය ගියේය. අද දක්වා නොනැසී පවතින විශේෂයන් උප පංති තුනක සංසන්දනාත්මක දුර්වල අවශේෂ නියෝජනය කරයි. උරගයින්ගේ වඳවීම පහසු වූයේ අහිතකර දේශගුණික විපර්යාස සහ ඇල්පයින් කඳු ගොඩනැගීමේ චක්රය සමඟ ඇති වූ වෘක්ෂලතා ආවරණයේ ස්වභාවයේ වෙනසක් පමණක් නොව, ඒ වන විට බරපතල තරඟකරුවන් බවට පත්ව තිබූ පක්ෂීන් සහ ක්ෂීරපායීන්ගේ දැඩි විශේෂණයෙනි. Mesozoic උරගයින්ගේ. කෙසේ වෙතත්, මෙම උසස් පෘෂ්ඨවංශීන්ගේ කාණ්ඩ දෙක බොහෝ කලකට පෙර ඔවුන්ගේ උරග මුතුන් මිත්තන්ගෙන් වෙන් වූ අතර, දිගු කලක් (වසර මිලියන දස දහස් ගණනක්) සංඛ්යාවෙන් ස්වල්පයක් වූ අතර, පෙනෙන විදිහට, සාපේක්ෂව රහසිගත ජීවන රටාවක් ගත කළේය.
ක්රිටේසියස් අවසානයේ පමණක් කුරුල්ලන් හා ක්ෂීරපායීන්ගේ වේගවත් සංවර්ධනය ආරම්භ වන අතර ඒ සමඟම උරගයින් වඳ වී යාම ආරම්භ වේ. පෘථිවියේ ගෝලීය වෙනස්කම් මගින් මෙය පහසු විය (කඳු ගොඩ නැගීමේ ක්රියාවලීන් සහ ගිනිකඳු ක්රියාකාරකම් වැඩි වීම, බොහෝ ප්රදේශවල මහාද්වීපික දේශගුණය වැඩි වීම යනාදිය; කොස්මික් සාධකවල බලපෑම පිළිබඳ උපකල්පන පවා සිදු කරන ලදී). පක්ෂීන් ( ඒව්ස්) ඉතා සංවිධිත උරගයන්ගෙන් වෙන් කර ඇත - සමහර archosaurs - පෙනෙන විදිහට ට්රයැසික් මධ්යයේ, වඩාත්ම පැරණි හා ප්රාථමික පක්ෂීන් දැන් දන්නා නමුත් ජුරාසික් යුගයේ (වසර මිලියන 135 ක් පමණ පැරණි) තැන්පතු වලින් පමණි. සමහර නවීන ඇණවුම්වල නියෝජිතයන් දැනටමත් ක්රිටේසියස් යුගයේ අගභාගයේ තැන්පතු වලින් සොයාගෙන ඇත.
ක්ෂීරපායින් ( ක්ෂීරපායී) කාලයාගේ ඇවෑමෙන් පැරණිතම උරග කණ්ඩායම් වලින් වෙන් විය - සතුන් වැනි උරගයින් (උප පන්තිය Theromorpha, seu උපාගම); උපාගම සමහර විට කාබනිෆරස් මධ්යයේ ඇති විය, පර්මියන් හි පුළුල් අනුවර්ණ විකිරණ අත්විඳ ඇති අතර ට්රයැසික් අවසානය වන විට අතුරුදහන් විය. එබැවින්, ක්ෂීරපායීන් සෑදීම, බොහෝ විට, ට්රයැසික් ආරම්භයේ දී (සමහර පාෂාණ විද්යාඥයින්ට අනුව, පර්මියන් අවසානයේ) සිදු විය යුතුය. Mesozoic ක්ෂීරපායීන්ගේ ඉතිහාසය දුර්වල ලෙස දන්නා කරුණකි. මෙසොසොයික් (ට්රයැසික්-ජුරාසික්) මධ්යයේ, සාපේක්ෂව ඉක්මනින් මිය ගිය කණ්ඩායම් කිහිපයක්, පෙනෙන විදිහට දැනටමත් වෙන් වී ඇත (නමුත් ඒවා අතර අපේ කාලය දක්වා නොනැසී පැවති මොනොට්රීම් ද විය). අඟහරු සහ වැදෑමහ ජුරාසික් වලින් ද සමහරක් ක්රිටේසියස් වලින් ද හැඳින්වේ. වැදෑමහ ක්ෂීරපායින්ගේ විස්තීර්ණ අනුවර්තන විකිරණ සහ නවීන ඇණවුම් ගොඩනැගීම දැනටමත් සෙනොසොයික් යුගයේ තෘතීයික කාල පරිච්ඡේදයේදී (වසර මිලියන 60-40 කට පමණ පෙර) සිදු විය.
එවැනි සංසන්දනය කිරීමෙන් චෝඩේට් වල අනුක්රමික පරිණාමය පිළිබඳ සංකේතාත්මක අදහසක් ලබා ගත හැකිය. පෘථිවි ග්රහලෝකයේ සමස්ත ඉතිහාසයේ පරිමාණය ලෙස අපි එක් පෘථිවි වර්ෂයක කාල පරිච්ඡේදයක් ගතහොත්, ජීවයේ මතුවීම මැයි මස අවසානයේදී - ජුනි මස මුලදී, පහළ වර්ගයේ අපෘෂ්ඨවංශීන්ගේ පෙනුම සිදුවනු ඇත. ජූනි මස අවසානය - ජූලි මස ආරම්භය, සහ අනෙකුත් අපෘෂ්ඨවංශීන් සහ වඩාත්ම ප්රාථමික චෝඩේට් - සැප්තැම්බර් අවසානයේ ( පැලියෝසොයික් යුගයේ කේම්බ්රියන් කාලය). ඔක්තෝබර් මැද භාගයේදී, පළමු පෘෂ්ඨවංශීන් දිස් වේ - ප්රාථමික හකු රහිත (ඕර්ඩොවිසියානු අවසානය - සිලුරියන් ආරම්භය), සහ ඔක්තෝබර් අවසානයේ (සිලූර්) පළමු හකු රහිත - ප්රාථමික මසුන් හකු රහිත අයගෙන් හුදකලා වේ. පළමු අවසානයේ - නොවැම්බර් දෙවන දශකයේ ආරම්භය (මැද ඩෙවෝනියන්), පළමු උභයජීවීන් lobe-finned මාළු වෙන්; බොහෝ විට, නොවැම්බර් අවසාන දින පහ ආරම්භයේදී (කාබනිෆරස් කාල පරිච්ඡේදයේ මැද භාගයේදී), පළමු උරගයන් දිස්වන අතර නොවැම්බර් අග සිට - දෙසැම්බර් පළමු දින (පර්මියන් කාලය), උභයජීවීන් වඳ වී යාම සහ උරගයින්ගේ මල් පිපීමේ ආරම්භය ආරම්භ වන අතර දෙසැම්බර් දෙවන දශකය අවසන් වන තෙක් (මුළු මෙසෝසොයික් යුගය) පවතී. ට්රයැසික් යුගයේ ආරම්භයේ දී (අපගේ පරිමාණයෙන් දෙසැම්බර් 3-4 දක්වා), පුරාණ ක්ෂීරපායින් ප්රාථමික උරගයන්ගෙන් වෙන් වූ අතර, එම කාලය අවසානයේ (දෙසැම්බර් 7-8), ප්රගතිශීලී උරගයන්ගෙන් වෙන් වූ පුරාණ පක්ෂීන් - ආචෝසෝරයන්.
කෙසේ වෙතත්, දෙසැම්බර් දෙවන දශකය අවසානයේ (ක්රිටේසියස් යුගයේ අවසානය) පමණක් කුරුල්ලන්ගේ හා ක්ෂීරපායින්ගේ වේගවත් සංවර්ධනය ආරම්භ වන අතර මෙසෝසොයික් උරගයින් බොහෝ කණ්ඩායම් වඳ වී යයි. සෙනොසොයික් යුගය - ඉහළ පෘෂ්ඨවංශීන්ගේ නවීන කණ්ඩායම් ගොඩනැගීමේ කාලය - දෙසැම්බර් 23 පමණ වන විට ආරම්භ වන අතර බොහෝ නවීන පවුල් ගොඩනැගීම - දෙසැම්බර් 28 සිට (නියෝජීන් ආරම්භය). ප්ලයිස්ටොසීන (චතුර්තික) යුගයේ ආරම්භය (මෙම පරිමාණයෙන්) දෙසැම්බර් 31 වන දින සවස 6-8 ට ආසන්න වශයෙන් වැටේ. ප්රාථමික (පැරණි) මිනිසුන් සහ නවීන හෝ නවීන ක්ෂීරපායින් සහ පක්ෂීන්ට සමීප වන කාලය මෙයයි. නූතන මිනිසා ( හෝමෝ සේපියන්ස්- සාධාරණ මිනිසා) පෙනී සිටියේ මීට වසර 100,000 කට පමණ පෙර, එනම්, අපගේ කාල පරිමාණයට අනුව, දෙසැම්බර් 31 වන දින අවසාන මිනිත්තු 20-15 තුළ පමණක් වන අතර, පුරාණ ඊජිප්තුවේ සිට අද දක්වා මානව සංස්කෘතියේ ඉතිහාසය ගත වන්නේ අවසාන 3 පමණි. - අවුරුද්දේ විනාඩි 5 යි!
chordates පරිණාමයේ කාල නිර්ණයපරිණාමීය ක්රියාවලියේ අසමානතාවය කෙරෙහි අවධානය යොමු කරයි, ප්රබල හැඩගැස්වීමේ කාල පරිච්ඡේද සාපේක්ෂව මන්දගාමී හා පටු අනුවර්තන පරිවර්තනයන් මගින් ප්රතිස්ථාපනය විය. A. N. Severtsov විසින් එවැනි නිත්ය ප්රත්යාවර්තයක් නම් කිරීමට යෝජනා කළේ ඇරෝමෝෆික් පරිණාමීය වෙනස්කම්වල කාල පරිච්ඡේදවල වෙනසක් ලෙස ජීවියාගේ අත්යවශ්ය ක්රියාකාරකම් ඉහළ ශක්ති මට්ටමකට (වැදගත් ක්රියාකාරකම්වල ශක්තියේ වැඩි වීමක්) ඉහළ නංවන රූප භෞතික විද්යාත්මක පරිවර්තනයන්ට මග පාදයි - මෝඩ අනුවර්තනය වීමේ කාල පරිච්ඡේද හෝ සංඛ්යාව වැඩි කිරීම, පුලුල්ව පැතිරීම, දේශීය තත්වයන්ට අනුවර්තනය වීම සහ යටත් කණ්ඩායම් (පවුල්, වර්ග, විශේෂ) වලට කැඩී යාමෙන් අත්පත් කරගත් වාසි සාක්ෂාත් කර ගැනීම. G. Osborne අවසාන ක්රියාවලිය අනුවර්තී විකිරණ ලෙස හැඳින්වේ. චෝඩේට් සෑදීම සහ පරිණාමය ඇරෝමෝෆික් මාර්ගයක සම්භාව්ය උදාහරණයකි - පන්ති මතුවීම - එක් එක් ඒවායේ පසුකාලීන මෝඩ අනුවර්තනය මල් පිපීම සමඟ, එමඟින් එක් එක් කාලවල සත්ත්ව විශේෂවල ප්රමුඛ ස්ථානයක් හිමි විය.
chordates පරිණාමයේ තවත් එක් අංගයක් කෙරෙහි අවධානය යොමු කළ යුතුය. Aromorphoses විසින් පිහිටුවන ලද නව පන්තියක්, සාමාන්යයෙන් එහි වර්ධනයේ මුල් අවධියේදී, යටත් ස්ථානයක් හිමි කර ගත් ශාඛා වෙන් කරන ලද අතර, බොහෝ විට අවාසි සහගත biotopes වෙත තල්ලු කරනු ලැබේ. නව පරිසරයන් ප්රගුණ කිරීමට සහ පිටසක්වල ජීවන තත්වයන්ට අනුවර්තනය වීමට බලකෙරුණු ඔවුන් සෙමෙන් පරිණාමය වූ නමුත් සාමාන්ය වැදගත්කමකින් යුත් අනුවර්තනයන් ලබා ගත හැකි අතර, ඒවා පිහිටුවීමෙන් සහ තහවුරු කිරීමෙන් පසු, ඔවුන්ගේ පූර්වගාමීන් බලහත්කාරයෙන් ඉවත් කිරීමට සිදුවිය. රීතියක් ලෙස, මෙයට පෙර පෘථිවි පෘෂ්ඨයේ (කඳු ගොඩ නැගීමේ චක්ර), දේශගුණය සහ වෘක්ෂලතා ආවරණය වෙනස් විය.
ඒ නිසා, අග්නාතාහා Gnathostomataපෙනෙන විදිහට, සමීප කාලයකදී පිහිටුවා ඇත, නමුත් පසුව හකු රහිත ඒවා, ගලා යන ජලයේ ජීවිතයට අනුවර්තනය වීම, හකු නොමැති අය බොහෝ පාරිසරික ස්ථානවලින් ඉවත් කිරීමට බල කෙරුනි. මසුන්ගේ පරිණාමය තුළද එයම සිදු විය: සන්නද්ධ මසුන් වෙනුවට කාටිලේජිනස් සහ දෙවැන්න අස්ථි මාළු මගින් ප්රතිස්ථාපනය විය. භෞමික පෘෂ්ඨවංශීන් (උභයජීවීන්) දර්ශනය වූ විට, ශාඛාවක් ඔවුන්ගේ පුරාණ කණ්ඩායමෙන් (ichthyostegs) කලින් වෙන් වූ අතර, එය පසුව උරගයන් බිහි වූ අතර, දෙවැන්න සෑදීමේ මුල් අවධියේදී, සත්ව-දත් සහිත සතුන් සමූහයක් වෙන් වී ඇති අතර, එය වර්ධනය විය. ක්ෂීරපායින් වෙත. නව පන්තියේ chordates ගොඩනැගීම සැමවිටම නව "අනුවර්තී කලාපයක්", නව වාසස්ථානයක් (G. Simpson) සංවර්ධනය කිරීම සමඟ සම්බන්ධ වී ඇති බව අවධාරණය කළ යුතුය. ඉතින්, මිරිදිය සඳහා මුහුද හැර ගිය chordates, පෘෂ්ඨවංශීන් බවට පත් විය; මත්ස්ය පංතිවල පරිණාමය සහ වෙනස්වීම් මෝය ආශ්රිත ජලයට සහ පහළ ප්රදේශයේ සිට ගංගාවල ඉහළ ප්රදේශ දක්වා අඛණ්ඩව විනිවිද යාම සමඟ සම්බන්ධ විය. ටෙට්රාපොඩ් (භෞමික) පෘෂ්ඨවංශීන්ගේ සුපිරි පන්තිය සඳහා මෙය වඩාත් පැහැදිලිය. පෘෂ්ඨවංශීන්ගේ තනි පන්ති විස්තර කිරීමේදී පරිණාමයේ කොන්දේසි, සාධක සහ මාර්ග වඩාත් විස්තරාත්මකව සලකා බලනු ලැබේ.
පෘෂ්ඨවංශීන්ගේ පරිණාමය තුළ, ව්යුහය (කායික සංවිධානය) පමණක් ක්රමානුකූලව වර්ධනය වූ නමුත්, ස්නායු පද්ධතියේ හා ක්රියාකාරිත්වයේ වර්ධනයේ පදනම මත පුද්ගලයන්ගේ සබඳතා වඩාත් සංකීර්ණ වූ අතර ජනගහන සංවිධානයේ වැදගත්කම වැඩි විය. සන්නිවේදනයේ සංකූලතාව යනු දෘශ්ය, ධ්වනි, රසායනික සහ වෙනත් නාලිකා හරහා සංකීර්ණ හා ධාරිතාවයෙන් යුත් තොරතුරු සම්ප්රේෂණය කිරීම කාර්යක්ෂම ප්රජනනය සහතික කිරීම, සතුන්ගේ අවකාශීය ව්යාප්තිය විධිමත් කිරීම, අභ්යවකාශයේ දිශානතිය වැඩි දියුණු කිරීම සහ පරිසරයට ඇති බලපෑම වැඩි දියුණු කිරීමයි. ජංගම කණ්ඩායම් (පවුල්, රංචු සහ රැළ) ස්වභාවික සම්පත් භාවිතා කිරීමේ හැකියාව පුළුල් කරන අතර පැවැත්ම සඳහා අරගලයේ අවස්ථා වැඩි කරයි.
යොමු කිරීම්: Naumov N.P., Kartashev N.N. පෘෂ්ඨවංශික සත්ව විද්යාව. - 2 කොටස. - උරගයන්, පක්ෂීන්, ක්ෂීරපායින්: ජීව විද්යාඥයින් සඳහා පෙළපොතක්. විශේෂඥ. විශ්ව විද්යාලය - එම්.: ඉහළ. පාසල, 1979. - 272 පි., අසනීප.
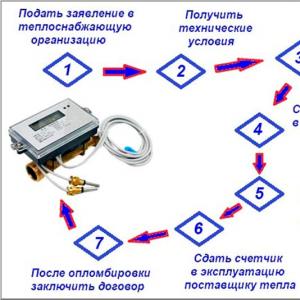
පෘෂ්ඨවංශික ඉන්ද්රිය පද්ධති
| පරාමිතිය නම | අර්ථය |
| ලිපි විෂය: | පෘෂ්ඨවංශික ඉන්ද්රිය පද්ධති |
| රුබ්රික් (තේමාත්මක කාණ්ඩය) | පරිසර විද්යාව |
| ඉන්ද්රිය පද්ධතිය | පද්ධති කාර්යයන් | පද්ධතියේ කොටසක් වන අවයව |
| සිරුරේ ආවරණ | ශරීරයේ අන්තර් ක්රියාකාරිත්වය සත්වයාගේ ශරීරය බාහිර පරිසරයෙන් වෙන් කරන අතර අහිතකර සාධකවල ක්රියාකාරිත්වයෙන් එය ආරක්ෂා කරයි. ඒ අතරම, සත්වයාගේ ශරීරයට අවශ්ය ද්රව්ය බාහිර පරිසරයට ලබා දීමට ඔවුන් ඉඩ නොදේ. බොහෝ සතුන් තුළ, ගෑස් හුවමාරු ක්රියාවලීන්හි ශරීරයේ සන්ධිය වැදගත් කාර්යභාරයක් ඉටු කරයි. | සම් |
| මස්කියුෙලොස්ෙකලටල් | සත්වයාගේ මස්කියුෙලොස්ෙකලටල් පද්ධතිය අභ්යවකාශයේ එහි චලනය සහතික කරයි. එය සත්වයාගේ ශරීරයේ රාමුවක් වන අතර එය යම් හැඩයක් පවත්වා ගැනීමට උපකාරී වේ. මස්කියුෙලොස්ෙකලටල් පද්ධතියේ තවත් කාර්යයක් ආරක්ෂිතයි - එය යාන්ත්රික හානිවලින් අභ්යන්තර අවයව ආරක්ෂා කරයි. | ඇටසැකිල්ල සහ මාංශ පේශි |
| ආහාර දිරවීමේ | ආහාර ජීර්ණ පද්ධතිය ශරීරයට පෝෂ්ය පදාර්ථ සපයයි. එය බාහිර පරිසරයෙන් ශරීරයට ඇතුළු වන කාබනික ද්රව්ය සකසයි. | දත්, දිව, කෙළ ග්රන්ථි, esophagus, ආමාශය, බඩවැල්, අක්මාව, අග්න්යාශය |
| ප්රවාහන | ප්රවාහන පද්ධතිය ශරීරයේ එක් කොටසක සිට තවත් ද්රව්යයකට වේගයෙන් මාරු වීම සහතික කරයි. එය හෝමෝන, පෝෂ්ය පදාර්ථ සහ පරිවෘත්තීය නිෂ්පාදන වාහකයක් වන බැවින් බොහෝ ක්රියාවලීන් නියාමනය කිරීමේදී එය වැදගත් කාර්යභාරයක් ඉටු කරයි. | හදවත, ධමනි, ශිරා, කේශනාලිකා |
| ශ්වසනය | ශ්වසන පද්ධතිය ශරීරයට ඔක්සිජන් ලබා දෙන අතර එයින් කාබන් ඩයොක්සයිඩ් ඉවත් කරයි. | පෙනහළු, උගුර, සම |
| පිටකිරීමේ | බැහැර කිරීමේ පද්ධතිය සත්වයාගේ ශරීරයෙන් පරිවෘත්තීය නිෂ්පාදන ඉවත් කරයි. | වකුගඩු, පෙනහළු, සම, මුත්රාශය, මුත්රාශය |
| අන්තරාසර්ග | අන්තරාසර්ග පද්ධතිය ශරීරයේ තනි අවයව හා ඉන්ද්රිය පද්ධතිවල ක්රියාකාරිත්වය නියාමනය කිරීමට සම්බන්ධ වේ. එය වෙනස්වන තත්වයන්ට ශරීරයේ දිගුකාලීන ප්රතික්රියා සපයයි. ජීව විද්යාත්මකව ක්රියාකාරී ද්රව්ය - හෝමෝන - අන්තරාසර්ග පද්ධතියේ සංඥා සම්ප්රේෂණය කිරීම සඳහා භාවිතා වේ. | හයිපොතලමස්, පිටියුටරි, අග්න්යාශය, ලිංගික ග්රන්ථි, තයිරොයිඩ්, පයිනල් ග්රන්ථිය |
| ස්නායු | ස්නායු පද්ධතිය සංවේද ඉන්ද්රියයන්ගේ ඇඟවීම් විකේතනය කරන අතර අනෙකුත් සියලුම අවයවවල ක්රියාවන් සමඟ සම්බන්ධීකරණය කරයි. එය ශරීරයේ තනි අවයව හා ඉන්ද්රිය පද්ධතිවල ක්රියාකාරිත්වය නියාමනය කිරීමට ද සම්බන්ධ වේ. ස්නායු පද්ධතිය වෙනස්වන තත්වයන්ට ශරීරයේ ඉක්මන් ප්රතික්රියා සපයයි | මොළය, සුෂුම්නාව, ස්නායු |
| ඉන්ද්රියයන් | සතුන්ගේ සංවේද ඉන්ද්රියයන් පරිසරයෙන් තොරතුරු වටහාගෙන ස්නායු පද්ධතියට සම්ප්රේෂණය කරයි. | ස්පර්ශ, සුවඳ, රසය, ඇසීම, පෙනීම, සමතුලිතතාවයේ අවයව |
| ප්රජනක | ප්රජනක පද්ධතිය සතුන්ගේ ප්රජනනය සහතික කරයි | ලිංගික අවයව |
3. බහු සෛලීය ජීවීන්ගේ කාර්යයන් නියාමනය කිරීම.
සතුන්ගේ ජීවියාගේ කාර්යයන් නියාමනය කිරීම පුද්ගලයෙකුගේ උදාහරණය මත වඩාත් හොඳින් සලකා බලනු ලැබේ. නියාමනය කිරීමේ මූලික ක්රම දෙකක් තිබේ: ස්නායු (ස්නායු සෛල පටල හරහා සම්ප්රේෂණය වන ස්නායු ආවේග භාවිතා කිරීම) සහ හාස්ය (විවිධ ශරීර තරල මගින් ගෙන යන රසායනික ද්රව්ය භාවිතා කිරීම).
හාස්ය නියාමනය- විවිධ ශරීර තරල (රුධිරය, වසා, පටක තරල) මගින් ගෙන යන රසායනික ද්රව්ය ආධාරයෙන් ශරීරයේ භෞතික විද්යාත්මක ක්රියාකාරකම් සම්බන්ධීකරණය කිරීම - හෝමෝන. අන්තරාසර්ග පද්ධතිය මගින් සිදු කරනු ලැබේ.
අන්තරාසර්ග පද්ධතිය- රුධිරයට හා වසා ගැටිති වලට හෝමෝන ස්රාවය කරන අවයව, අවයවවල කොටස් සහ තනි සෛල කට්ටලයක්. ස්නායු පද්ධතිය සමඟ එක්ව, එය මිනිස් සිරුරේ වැදගත් කාර්යයන් නියාමනය හා සම්බන්ධීකරණය කරයි: වර්ධනය, ප්රතිනිෂ්පාදනය, පරිවෘත්තීය, අනුවර්තනය කිරීමේ ක්රියාවලීන්.
අන්තරාසර්ග පද්ධතිය තුළ, මධ්යම සහ පර්යන්ත කොටස් වෙන්කර හඳුනාගෙන ඇති අතර, ඒවා එකිනෙකා සමඟ අන්තර් ක්රියාකාරී වන අතර තනි පද්ධතියක් සාදයි. අන්තරාසර්ග පද්ධතියේ ඵලදායී ක්රියාකාරිත්වය සඳහා පදනම වන්නේ ප්රතිපෝෂණ මූලධර්මය භාවිතා කිරීමයි.
මිනිස් සිරුරේ ක්රියාවලීන්ගේ ස්නායු නියාමනය සිදු කරනු ලබන්නේ සොමාටික් සහ ස්වයංක්රීය ස්නායු පද්ධතිවල ආධාරයෙන් ය.
සෝමාටික් ස්නායු පද්ධතිය සමන්විත වන්නේ අස්ථි මාංශ පේශි සහ සංවේදී ඉන්ද්රියයන් නවීකරණය කරන මධ්යම හා පර්යන්ත ස්නායු පද්ධතියේ කොටස් වලින් ය. එය ශරීරයට බාහිර පරිසරයෙන් තොරතුරු දැනගැනීම මෙන්ම බාහිර සාධකවලට ප්රතිචාර වශයෙන් ක්රියා (අස්ථි මාංශ පේශිවල විවිධ චලනයන් ආකාරයෙන්) සපයයි.
සොමාටික් ස්නායු පද්ධතිය මගින් සපයනු ලබන චලනයන් ක්රියාත්මක කරනු ලබන්නේ තනි මෝටර් ඒකකවල සම්බන්ධීකරණ ක්රියාවන් භාවිතා කරමිනි (මාංශ පේශි තන්තු කණ්ඩායම්, ඒ සෑම එකක්ම එක් මෝටර් නියුරෝනයකින් නවීකරණය කර ඇත).
ස්වයංක්රීය (ශාකමය) ස්නායු පද්ධතිය- අභ්යන්තර අවයව, ග්රන්ථි, රුධිර වාහිනී, සිනිඳු සහ සමහර ඉරි සහිත මාංශ පේශිවල ක්රියාකාරිත්වය නියාමනය කරන ස්නායු පද්ධතියේ කොටසක් වන අතර පරිවෘත්තීය ක්රියාවලීන් ද කළමනාකරණය කරයි.
ස්වයංක්රීය ස්නායු පද්ධතිය ශරීරයේ අවයව හා පටක වලට ප්රතිවිරුද්ධ බලපෑමක් ඇති කොටස් දෙකකින් සමන්විත වේ - සානුකම්පිත සහ පැරසිම්පතටික් බෙදීම්. ස්වයංක්රීය ස්නායු පද්ධතියේ ඉහළම පාලන මධ්යස්ථානය වන්නේ හයිපොතලමස් වන අතර එය අන්තරාසර්ග පද්ධතියේ ක්රියාකාරිත්වය ද පාලනය කරයි.
ස්වයංක්රීය ස්නායු පද්ධතිය අභ්යන්තර අවයව, සනාල පද්ධතිය, ග්රන්ථි සහ සිනිඳු මාංශ පේශි සඳහා නවෝත්පාදනයක් සපයයි. එය අස්ථි මාංශ පේශි මත ද trophic බලපෑමක් ඇත. මෙම මාංශ පේශි හැකිලීමකින් තොරව, එය ඔවුන්ගේ පෝෂණය වැඩි දියුණු කරන අතර එමඟින් ඔවුන්ගේ වැඩ උත්තේජනය කරයි. එය අභ්යන්තර අවයව හා රුධිර නාල වල ක්රියාකාරිත්වය, ග්රන්ථි වල ස්රාවය සහ හදවතේ ක්රියාකාරිත්වය නියාමනය කරයි. පරිවෘත්තීය ක්රියාවලීන් ස්වයංක්රීය ස්නායු පද්ධතිය මගින් ද නියාමනය කරනු ලැබේ.
ස්වයංක්රීය ස්නායු පද්ධතියේ ක්රියාකාරිත්වය මිනිසාගේ කැමැත්තට සහ විඥානයට යටත් නොවේ. පුද්ගලයෙකුට බොහෝ අභ්යන්තර අවයව පවතින බවක් දැනෙන්නේ නැත, විශේෂයෙන් චලනය නොවන ඒවා, උදාහරණයක් ලෙස ග්රන්ථි, ඒවා ස්රාවය වන ආකාරය, බඩවැල්වල ආහාර අවශෝෂණය වන ආකාරය සහ ඒ හා සමාන බවක් දැනෙන්නේ නැත. පුද්ගලයෙකුට ඔහුගේ මාංශ පේශි පාලනය කරන බැවින්, මෙම අවයවවල ක්රියාකාරිත්වය දැනුවත්ව පාලනය කළ නොහැකිය. එවැනි ක්රියාවලීන් පුද්ගලයෙකුගේ විඥානයෙන් පිටත සිදු වන අතර ඇයගේ කැමැත්තට යටත් නොවේ.
ස්වයංක්රීය ස්නායු පද්ධතියේ, සොමැටික් මෙන්, මධ්යම සහ පර්යන්ත කොටස් ඇත. මධ්යම කොටස වෘක්ෂලතා නියුරෝන මගින් නියෝජනය වන අතර, මොළයේ හා කොඳු ඇට පෙළේ සමුච්චය සාදයි - ශාකමය න්යෂ්ටීන්. පර්යන්ත කොටස සෑදී ඇත්තේ බොහෝ ස්වයංක්රීය ස්නායු නෝඩ් සහ ස්නායු තන්තු මගිනි.
පෘෂ්ඨවංශික ඉන්ද්රිය පද්ධති - සංකල්පය සහ වර්ග. "පෘෂ්ඨවංශීන්ගේ අවයව පද්ධති" කාණ්ඩයේ වර්ගීකරණය සහ විශේෂාංග 2017, 2018.
පෘෂ්ඨවංශික සතුන්ගේ සංවිධානය
පෘෂ්ඨවංශීන් පොදු morphophysiological සංවිධානයක් මගින් එක්සත් වේ. මෙම සතුන්ගේ සියලුම ඉන්ද්රිය පද්ධතිවල, සමජාතීය අවයවවල පරිණාමීය පරිවර්තනය හා සම්බන්ධ අනුක්රමික වෙනස්කම්වල ලක්ෂණ සොයාගත හැකිය. පහත දැක්වෙන්නේ තනි ඉන්ද්රිය පද්ධතිවල ව්යුහය, ක්රියාකාරිත්වය සහ ඔන්ටොජෙනිය තුළ තැබීම පිළිබඳ සාමාන්ය සැලැස්මකි.
සමඉතා වැදගත් ක්රියාකාරී පද්ධතියක් නියෝජනය කරයි. සම බාහිර පරිසරය සමඟ සෘජුව සම්බන්ධ වන අතර එය සෘජුවම බලපායි. සම සහ මාංශ පේශි මතුපිට සිට සත්වයාගේ ශරීරය සාදයි, එයට හැඩය ලබා දෙන අතර සියලුම අභ්යන්තර අවයව රඳවා තබා ගනී. සමේ ආවරණ බාහිර යාන්ත්රික හා රසායනික හානි, උෂ්ණත්වයට නිරාවරණය වීම, වියළීම, ක්ෂුද්ර ජීවීන්ගේ විනිවිද යාමෙන් ශරීරය ආරක්ෂා කරයි. සම තාපගතිකරණය, ගෑස් හුවමාරුව සහ දිරාපත්වන නිෂ්පාදන බැහැර කිරීම සඳහා සහභාගී වේ. සමේ ව්යුත්පන්නයන් චලනය වීමේ අවයව (කුර), ග්රහණය කර ගැනීම (නියපොතු), ප්රහාර සහ ආරක්ෂාව (අං, ඉඳිකටු, ආදිය), පියාසර කිරීම (නැමීම්), පිහිනීම (වෙබ්) සඳහා සේවය කළ හැකිය.
සමේ ස්පර්ශයේ අවයවවල ප්රතිග්රාහක අඩංගු වේ, එය විවිධ අරමුණු සඳහා බොහෝ ග්රන්ථි ඇත (ශ්ලේෂ්මල, මේද, ගන්ධය, දහඩිය, ආදිය).
පෘෂ්ඨවංශික සම ස්ථර දෙකකින් සංලක්ෂිත වේ. පිටත තට්ටුව - එපීඩර්මිස් - ectodermal සම්භවයක් ඇත. එය සෑම විටම බහු ස්ථර වේ. එහි පහළ ස්ථරය ජීවය සඳහා ජීවමානව හා ක්රියාකාරීව පවතින අතර නව සෛල ස්ථර නිපදවයි. එපීඩර්මිස් හි ඉහළ ස්ථර සාමාන්යයෙන් සමතලා වූ සෛල වලින් සමන්විත වන අතර, භූමිෂ්ඨ පෘෂ්ඨවංශීන් තුළ කෙරටිනීකරණය වී, මිය යන අතර නිරන්තරයෙන් මිය යයි.
මන්දගාමී වීම. එපීඩර්මිස් සමේ අං ව්යුත්පන්නයන් ඇති කරයි - අං කොරපොතු, පිහාටු, හිසකෙස්, නියපොතු, කුර, හිස් අං. අපිචර්මයේ විවිධ සමේ ග්රන්ථි වර්ධනය වේ.
සමේ අභ්යන්තර තන්තුමය ස්ථරය කොරියම්, වෙනත් ආකාරයකින් සඳහන් වේ cutis, හෝ සම ම, mesodermal rudiment සිට වර්ධනය වේ - somite හි සමේ පත්රය. කෝරියම් තට්ටුව ඝනයි, එය විශාල ශක්තියක් ඇති සමේ ප්රධාන කොටස සෑදෙයි. කෝරියම් තුළ, විවිධ අස්ථිගත කිරීම් මාළු කොරපොතු, අන්තර් අස්ථි, ඊනියා සමේ ඇටසැකිල්ල (කොන්ඩ්රල් ඇටසැකිල්ලට ප්රතිවිරුද්ධව) සාදයි. සමේ වියදමින් මුවන්ගේ අස්ථි අං ද වර්ධනය වේ. කපනයෙහි පහළ කොටසෙහි, මේදය චර්මාභ්යන්තර ස්ථරයක් එකතු වේ.
මාංශ පේශී. සමට යටින් පිහිටා ඇති මාංශ පේශී ස්තරය යනු මාංශ පේශිවල විශාල කොටසක් ලෙස හැඳින්වේ ශරීරයේ මාංශ පේශි, හෝ ශාරීරික. එය සතුන්ට පරිසරයේ චලනය වීමේ හැකියාව ලබා දෙන අතර ඉරි සහිත මාංශ පේශි පටක වලින් සමන්විත වේ. පහළ පෘෂ්ඨවංශීන් තුළ, හිස්කබල නොවන අයගේ මෙන්, මාංශපේශී ඛණ්ඩිත චරිතයක් ඇත. ඉහළ පෘෂ්ඨවංශීන් තුළ, ශරීරයේ චලනයන්හි සාමාන්ය සංකූලතාව හේතුවෙන්, අත් පා වර්ධනය වීමත් සමඟ, ඛණ්ඩනය බාධා ඇති වන අතර, කඳ මාංශ පේශි කාණ්ඩගත වී, කඳ, හිස සහ චලනය වන අවයව වැනි ශරීරයේ කොටස් සාදයි.
හැර ශාරීරික මාංශ පේශිපෘෂ්ඨවංශීන්ට බඩවැල්වල මාංශ පේශී සහ අනෙකුත් අභ්යන්තර අවයව (නාල, ඇල) ඇත. මෙම මාංශ පේශි visceral ලෙස හැඳින්වේ. එය සිනිඳු මාංශ පේශි පටක වලින් සමන්විත වන අතර, විශේෂයෙන්, බඩවැල්වල ආහාර චලනය, රුධිර නාල වල බිත්ති හැකිලීම සපයයි.
කඳේ මාංශපේශී කළල ලෙස මයෝටෝමයේ අභ්යන්තර ස්ථරයෙන් පැන නගී (ලැන්ස්ලට් වර්ධනය බලන්න), i.e. පෘෂ්ඨීය mesoderm. visceral musculature යනු පාර්ශ්වීය තහඩුවේ ව්යුත්පන්නයකි, i.e. උදරීය මෙසෝඩර්ම්.
අභ්යන්තර ඇටසැකිල්ල- පෘෂ්ඨවංශික සතෙකුගේ ශරීරයේ ආධාරක පදනම මෙයයි. මීට අමතරව, ඇටසැකිල්ල ශරීරයේ චලනය සම්බන්ධ වේ, අභ්යන්තර අවයව ආරක්ෂා කරයි.
භූගෝලීය වශයෙන්, පෘෂ්ඨවංශීන්ගේ ඇටසැකිල්ල අක්ෂීය, අභ්යන්තර, අත් පා පටිය සහ නිදහස් අත් පා ඇටසැකිලි ලෙස බෙදිය හැකිය.
එහි මුල් ස්වරූපයෙන් අක්ෂීය ඇටසැකිල්ල ඉදිරිපත් කර ඇත ස්වරයඝන සම්බන්ධක පටක පටලයකින් වට වී ඇත. දෙවැන්න ස්වරය පමණක් නොව, ඊට ඉහලින් පිහිටා ඇති ස්නායු නාලය ද ආවරණය කරයි. ප්රාථමික අන්ත්රයේ පෘෂ්ඨීය පැත්තේ ප්රාථමිකයෙන් නෝටෝකෝඩ් වර්ධනය වේ, i.e. අන්තරාසර්ග සම්භවයක් ඇත.
සහල්. 9. කශේරුකාවක් සෑදීම (තීර්යක් කොටස්):
1 - පහළ චාපයේ ආරම්භය; 2 - ඉහළ චාපයේ ආරම්භය; 3 - ඉළ ඇටය, 4 - හේමාල් ක්රියාවලිය, 5 - කශේරුකාවේ පාර්ශ්වීය ක්රියාවලිය, 6 - උසස් ආරුක්කු
බොහෝ පෘෂ්ඨවංශීන් තුළ, නොටෝකෝඩ් විස්ථාපනය කර කාටිලේජීය හෝ අස්ථි ඇටසැකිල්ලක් මගින් ප්රතිස්ථාපනය වේ. කාටිලේජීය සහ අස්ථි ඇටසැකිලි ඉහත සඳහන් කළ සම්බන්ධක පටක (මෙසෝඩර්ම් සම්භවය) පටලයේ ව්යුත්පන්නයන් ලෙස වර්ධනය වේ. මෙම කවචය මෙලෙස ඇටසැකිල්ලකි.
අක්ෂීය ඇටසැකිල්ලේ ඇත පෘෂ්ඨවංශික තීරුව සහ මස්තිෂ්ක හිස් කබල. කශේරුකා වර්ධනය අතරතුර, නොටෝකෝඩ් මතුපිටට යාබදව පාරමිතිකව පිහිටා ඇති යුගල කාටිලේජ මුලින් තබා ඇත. මේවා ඉහළ සහ පහළ පෘෂ්ඨවංශික ආරුක්කු වල මූලයන් වේ (රූපය 9.1). ඉහළ ආරුක්කු වල පිටත කෙළවරේ වර්ධනය හා වසා දැමීම ගොඩනැගීමට හේතු වේ කොඳු ඇට පෙළ, ස්නායු නාලය පිහිටා ඇති (රූපය 9, II). පහළ චාප කෞඩල් කලාපයේ (මාළු වල) වසා දමා සීමාව හේමල් ඇල,පෘෂ්ඨීය ධමනිය සහ වලිග නහරය ගමන් කරන්නේ කොතැනින්ද? ඉහළ සහ පහළ ආරුක්කු වල අභ්යන්තර කෙළවර වසා දැමීමේ ප්රතිඵලයක් වශයෙන්, පෘෂ්ඨවංශික සිරුරු සෑදී ඇත, ඇතුළත සහ ඒවා අතර, යම් දුරකට, notochord සංරක්ෂණය කළ හැකිය (රූපය 9, III). කඳ කලාපයේ, පහළ පෘෂ්ඨවංශික ආරුක්කු වල ක්රියාවලීන් සඳහා ඉළ ඇට බැඳී ඇත.
මොළයේ හිස් කබල, හෝ හිස් කබල, මොළයේ මූලද්රව්යයට යටින් කාටිලේජ යුගල දෙකක ස්වරූපයෙන් තැන්පත් කර ඇත. ඔවුන්ගේ පසුපස යුගලය - පැරචෝඩලියාව- ෙකෝඩ් ඉදිරිපස කෙළවරේ පැතිවල පිහිටා ඇත; ඉදිරිපස යුගල - trabeculae- ඇය ඉදිරිපිට. ඔවුන් පළමු කශේරුකාවේ මුලික කොටස් සහ අර්ධ වශයෙන් අභ්යන්තර ඇටසැකිල්ලේ ආරුක්කු පරිවර්තනය කරයි. පැරචෝර්ඩලියා සහ ට්රැබෙකියුලේ වර්ධනය හා වසා දැමීම මොළයට යටින් ඇති හිස් කබලේ ප්රධාන ලැමිනා සෑදීමට හේතු වේ. ඒ සමගම, කාටිලේජීය කැප්සියුල සෑදෙන සංවේදී ඉන්ද්රියයන් (සුවඳ, පෙනීම, ශ්රවණය) වටා දිස් වේ. ඒවා ප්රධාන මට්ටමට වඩා තරමක් ඉහළින් පිහිටා ඇත

සහල්. 10. මෝර කාටිලේජිනස් හිස් කබල වර්ධනය:
1 - ගිල් ආරුක්කු; 2 - හයියිඩ් ආරුක්කු; 3 - හකු ආරුක්කු; 4 - ශ්රවණ කැප්සියුලය; 5 - ස්වරය; 6 - පිටියුටරි ග්රන්ථිය; 7 - බඩවැල්; 8 - මැද මොළය; 9 - කොඳු ඇට පෙළ; 10 - නාස්පුඩුව; 11 - ඇස; 12 - පැරචෝර්ඩලියා; 13 - ප්රාථමික පූර්ව මොළය; 14-කක්ෂීය කාටිලේජ; 15 - trabeculae
හිස් කබලේ තහඩු සහ පැතිවලින් මොළය ආවරණය කරයි. පසුකාලීන වර්ධනයේ දී, සංවේදී ඉන්ද්රියන්ගේ කැප්සියුල හිස්කබලේ ප්රධාන ලැමිනා සමඟ සම්බන්ධ වී ඒකාබද්ධ වේ. හිස් කබලේ කාටිලේජ තත්වයේදී, මොළයේ ආවරණයේදී සම්පූර්ණ වහලක් ඇති නොවේ. හිස් කබලේ වහලයේ ලින්ටල් අතර ඉතිරිව ඇති සිදුරු - ෆොන්ටනෙල්ස්- සම්බන්ධක පටක පටලයකින් තද කර ඇත. හිස්කබලේ ඝන වහලක් සිදු වන්නේ අධිස්ථාපිත (සමේ) අස්ථි (ඉදිරිපස, ප්රාචීරය) සෑදීම සම්බන්ධව පමණි.
මේ අනුව, මොළයේ හිස් කබල ඔවුන්ගේ ආරක්ෂිත ගොඩනැගීම ලෙස මොළයේ හා සංවේදී ඉන්ද්රියන්ගේ වර්ධනය සම්බන්ධව පැන නගී.
දෘශ්ය ඇටසැකිල්ල phylogenetically හිස් කබලෙන් ස්වාධීනව පිහිටුවා ඇත (රූපය 10). එහි තැබීම සිදු වන්නේ ආහාර දිරවීමේ නලයේ ඉදිරිපස අසල ඇති සම්බන්ධක පටක වලය. මුලදී, visceral ඇටසැකිල්ල යනු gill slits අතර පිහිටා ඇති බොහෝ ඒකාකාරී ආරුක්කු මාලාවකි. ඔවුන් ශ්වසන උපකරණ සඳහා ආධාරකයක් ලෙස සේවය කරයි.
ඉන්ද්රිය ඇටසැකිල්ලේ පසුකාලීන පරිවර්තනය ඉහළ සහ පහළ ප්රාථමික හකු, මැද කණ, මස්තිෂ්ක හිස් කබලේ පතුල සහ ස්වරාලය වැනි අවයව පෘෂ්ඨවංශීන් විසින් අත්පත් කර ගැනීම සමඟ සම්බන්ධ වේ.
"අභ්යන්තර ඇටසැකිල්ල" යන සංකල්පය ප්රධාන වශයෙන් පහත් පෘෂ්ඨවංශීන් සම්බන්ධයෙන් සැලකේ. උසස් අධ්යාපනයේ දී එය "අභ්යන්තර හිස් කබල", "මුහුණු හිස් කබල" යන සංකල්ප මගින් ප්රතිස්ථාපනය වේ.
අත් පා පටිය සහ නිදහස් අත් පා වල ඇටසැකිල්ල. පෘෂ්ඨවංශීන් තුළ, යුගල නොකළ සහ යුගල කරන ලද අත් පා වෙන්කර හඳුනාගත හැකිය. අනෙක් අතට, යුගල කරන ලද අත් පා වරල් හෝ බිම් ආකාරයේ අත් පා විය හැකිය.
යුගල නොකළ අත් පා ඇටසැකිල්ල - පෘෂ්ඨීය, කෞඩල්, ගුද වරල්- ඇටසැකිල්ලේ අනෙකුත් කොටස් සමඟ සම්බන්ධ නොවන කාටිලේජීය හෝ අස්ථි කිරණ ගණනාවකින් සමන්විත වේ.
යුගල වූ අත් පා වල ඇටසැකිල්ල අත් පා පටි වල ඇටසැකිල්ල සහ නිදහස් අවයවයේ ඇටසැකිල්ල ලෙස බෙදා ඇත. අත් පා පටි සෑම විටම සත්වයාගේ ශරීරය තුළ පිහිටා ඇත. පෘෂ්ඨවංශීන්ගේ නිදහස් අත් පා ඇටසැකිල්ල වර්ග දෙකකි: මාළු වරල් සහ

සහල්. 11. පෘෂ්ඨවංශික ඇටසැකිලි යෝජනා ක්රමය:
I - මාළු ඇටසැකිල්ල, II - භූමිෂ්ඨ පෘෂ්ඨවංශික ඇටසැකිල්ල
ඇඟිලි පහක් සහිත අවයවයභූමිෂ්ඨ පෘෂ්ඨවංශීන්. පළමු අවස්ථාවේ දී, ඇටසැකිල්ල තනි ලීවරයක් ලෙස පටියට සාපේක්ෂව චලනය වන කාටිලේජ හෝ අස්ථි පේළි කිහිපයකින් නියෝජනය වේ. ඇඟිලි පහක් සහිත පාදයක ඇටසැකිල්ල, ලිවර්ස් ගණනාවකින් සමන්විත වන අතර එය අත් පා පටියට සාපේක්ෂව එකට ගමන් කළ හැකි අතර වෙන වෙනම - එකක් අනෙකට සාපේක්ෂව (රූපය 11). අත් පා ඇටසැකිල්ල තැබීම සමේ සම්බන්ධක පටක ස්ථරයේ සිදු වේ.
ආහාර ජීර්ණ අවයව. ආහාර ජීර්ණ පද්ධතිය මුඛයෙන් ආරම්භ වී ගුද මාර්ගයෙන් අවසන් වන නලයක් මගින් නිරූපණය කෙරේ. ආහාර ජීර්ණ පත්රිකාව සෑදී ඇත්තේ ගැස්ට්රුලයේ එන්ඩොඩර්මල් නළයෙනි (ලැන්ස්ලට් වර්ධනය බලන්න). මේ සම්බන්ධයෙන්, ආහාර ජීර්ණ පත්රයේ එපිටිලියම් එන්ඩොඩර්මල් වේ. එන්ඩොඩර්මල් එපිටිලියම් නොපෙනෙන ලෙස ectodermal වෙත ගමන් කරන්නේ මුඛ සහ ගුද විවරයන් ඇති කලාපය තුළ පමණි. මෙය ඉහත සිදුරු සෑදීමේදී ශරීරයේ බිත්ති (සහ, ප්රතිඵලයක් වශයෙන්, ectoderm) ආක්රමණය කිරීම නිසාය.
ආහාර ජීර්ණ පත්රිකාව පහත සඳහන් ප්රධාන කොටස් වලට බෙදා ඇත: 1) මුඛ කුහරයආහාර ගැනීම සඳහා සේවය කිරීම; 2) ෆරින්ක්ස්- සෑම විටම ශ්වසන ඉන්ද්රියයන් සමඟ සම්බන්ධ වූ දෙපාර්තමේන්තුවක්: මාළු, ෆරින්ක්ස් තුළට ගිල් ස්ලිට් විවෘත වේ, භූමිෂ්ඨ පෘෂ්ඨවංශීන් තුළ, ෆරින්ක්ස් හි ස්වරාලය ස්ලිට් පිහිටා ඇත; ෆරින්ක්ස් නිවැරදිව ආහාර ජීර්ණ නාලයේ ශ්වසන අංශය ලෙස හැඳින්වේ; 3) esophagus, 4) බඩ- සමහර අවස්ථාවලදී ඉතා සංකීර්ණ උපාංගයක් ඇති අන්ත්ර පත්රිකාව පුළුල් කිරීම; 5) බඩවැල්, සාමාන්යයෙන් ඉදිරිපස, හෝ තුනී, මැද, හෝ ඝන, සහ පසුපස හෝ ගුද මාර්ගයට බෙදා ඇත. පෘෂ්ඨවංශීන් මාලාවක ආන්ත්රික පත්රිකාවේ රූප විද්යාත්මක සංකූලතාව මාර්ගය අනුගමනය කරයි
එහි දිගු කිරීම සහ දෙපාර්තමේන්තු වලට වෙන් කිරීම. ආහාර දිරවීමේ ග්රන්ථි වර්ග තුනක නාල ආහාර දිරවීමේ නලයට විවෘත වේ: කෙල, අක්මාව, අග්න්යාශය.
ලවණ ග්රන්ථි- භූමිෂ්ඨ පෘෂ්ඨවංශීන් අත්පත් කර ගැනීම. ඔවුන් මුඛ කුහරයේ ශ්ලේෂ්මල ග්රන්ථි පරිවර්තනය කරයි. ඔවුන්ගේ රහස ආහාර තෙතමනය හා කාබෝහයිඩ්රේට බිඳවැටීම ප්රවර්ධනය කරයි.
අක්මාව සහ අග්න්යාශය වර්ධනය වන්නේ කලල බඩවැල්වල ඉදිරිපස කොටස ඉදිරියට නෙරා ඒමෙනි. අක්මාව හටගන්නේ අන්ත්රයේ උදර බිත්තියේ අන්ධ වර්ධනයක් මගිනි (ලැන්ස්ලට් බඩවැලේ රක්තපාත වර්ධනය බලන්න). අක්මා නාලිකා ඉදිරිපස කුඩා අන්ත්රය තුළට ගලා යයි. අග්න්යාශයකිහිප දෙනෙකුගෙන්, බොහෝ විට තුනකින්, මුලික කොටස් වලින් වර්ධනය වේ, ඒවා ද බඩවැලේ වර්ධනය වේ. මෙම ග්රන්ථිය, අක්මාව මෙන් නොව, සාමාන්යයෙන් සංයුක්ත සිරුරේ පෙනුමක් නොමැති අතර, එහි lobules ඉදිරිපස කුඩා අන්ත්රයේ mesentery පුරා විසිරී ඇත.
මෙම ග්රන්ථි දෙකේම ක්රියාකාරීත්වය ආහාර දිරවීමට වඩා පුළුල් ය. ඉතින්, අක්මාව, පිත ස්රාවය කිරීම, මේද ඉමල්සිෆයි කිරීම සහ අනෙකුත් ආහාර ජීර්ණ එන්සයිම වල ක්රියාකාරිත්වය සක්රීය කිරීමට අමතරව වැදගත් පරිවෘත්තීය ඉන්ද්රියයක් ලෙස සේවය කරයි. මෙන්න, සමහර හානිකර දිරාපත් නිෂ්පාදන උදාසීන, glycogen සමුච්චය. අග්න්යාශයික එන්සයිම ප්රෝටීන, මේද සහ කාබෝහයිඩ්රේට් බිඳ දමයි. ඒ අතරම, අග්න්යාශය අභ්යන්තර ස්රාවයේ අවයවයක් ලෙස සේවය කරයි. මෙම කාර්යයේ බිඳවැටීම ශරීරයේ සීනි භාවිතා කිරීමට ඇති හැකියාව අහිමි වීමට හේතු වේ. ප්රතිඵලය දරුණු රෝගයකි - දියවැඩියාව.
ශ්වසන පද්ධතියපෘෂ්ඨවංශීන් වර්ග දෙකක් ඇත - ගිල් සහ පෙනහළු, සහ පෘෂ්ඨවංශීන්ගෙන් සැලකිය යුතු කොටසක, ශ්වසනයේදී සම අත්යවශ්ය වේ.
ගිල් උපකරණ යනු යුගලනය කරන ලද, සාමාන්යයෙන් සමමිතිකව පිහිටා ඇති, ෆරින්ක්ස් බාහිර පරිසරය සමඟ සන්නිවේදනය කිරීමට සේවය කරන ස්ලිට් පද්ධතියකි. ජිල් ස්ලිට් වල ඉදිරිපස සහ පසුපස බිත්ති ශ්ලේෂ්මල පටලයකින් පෙලගැසී ඇති අතර එය ලැමිලර් පිටතට වර්ධනය වේ; වර්ධනය බෙදී ඇත පෙතිනම දරන ගිල්. පෙති වලට ඉහළින් ඇති සෑම ගිල් තහඩුවක්ම නම් කර ඇත අර්ධ-ගිලි. ගිල් ස්ලිට් අතර පරතරයන් (ගිල් සෙප්ටා තුළ) දිරාපත් වේ visceral gill arches(අභ්යන්තර ඇටසැකිල්ල සඳහා 27 පිටුව බලන්න). මේ අනුව, එක් එක් ගිල් ආරුක්කු එකිනෙකට වෙනස් ගිල් ස්ලිට් දෙකක අර්ධ-ගිල් දෙකකට සම්බන්ධ වේ.
ගිල් ස්ලිට්ස්ෆරින්ක්ස් සිට පිටතට වැඩෙන යුගල එන්ඩෝඩර්මල් නෙරා යාමේ පද්ධතියක ස්වරූපයෙන් තැන්පත් කර ඇත. ඒ සමගම, පිටත ඉන්ටෙග්මන්ට් වල ectodermal invaginations පෙනේ. primordia එකිනෙකා දෙසට වර්ධනය වී පසුව එක්සත් වේ. එමනිසා, ගිල් ස්ලිට්ස් මිශ්ර එන්ටෝ සහ චර්ම-චර්ම සම්භවයක් ඇත. Gill සූතිකා සාමාන්යයෙන්
විඛණ්ඩනයේ ectodermal මූලයෙන් වර්ධනය වන අතර, හකු රහිත සතුන් තුළ පමණක් ඔවුන්ට අන්තරාසර්ග සම්භවයක් ඇත.
භූමිෂ්ඨ පෘෂ්ඨවංශීන්ගේ ශ්වසන අවයව - පෙනහළු- යෝජනා ක්රමයේ ඒවා ස්වරාල විඛණ්ඩනය හරහා ෆරින්ක්ස් තුළට විවෘත වන බෑග් යුගලයකි. කලලරූපීව, පෙනහළු පැන නගින්නේ ගිල් උපකරණයේ පිටුපස ඇති ෆරින්ක්ස් හි උදර බිත්තියේ නෙරා යාමේ ස්වරූපයෙන් ය. අන්තරාසර්ග සම්භවයක් ඇත. කලල විකසනයේ මුල් අවධියේදී පෙනහළු අංකුර අභ්යන්තර (එන්ඩෝඩර්මල්) ජිල් ස්ලිට් යුගලයකට සමාන වේ. මෙම තත්ත්වයන් මෙන්ම පෙනහළුවලට සහ ජිල් වලට පොදු වූ රුධිර සැපයුමේ සහ නවෝත්පාදනයේ ලක්ෂණ, පෙනහළු පසුපස ජිල් මලු යුගලයේ සමජාතීය ලෙස සැලකීමට අපට බල කරයි.
සම්ඝන අං හෝ අස්ථි කොරපොතු නොමැති අවස්ථාවන්හිදී ශ්වසනයට සහභාගී වේ, නිදසුනක් ලෙස, උභයජීවීන්, නිරුවත් සමක් ඇති මාළු.
ක්රියාකාරී ලෙස, ශ්වසන පද්ධතිය ඔක්සිජන් සමඟ රුධිරය පොහොසත් කිරීම සහ කාබන් ඩයොක්සයිඩ් ඉවත් කිරීම සම්බන්ධ වේ. පහළ ජලජ සතුන්ගේ ශ්වසන පද්ධතිය හරහා ඇමෝනියා නිකුත් වේ. උණුසුම්-ලේ සහිත සතුන් තුළ, එය තාපගතිකරණයේ ක්රියාවලීන් සඳහා සම්බන්ධ වේ. ශ්වසන පද්ධතියේ ක්රියාකාරිත්වයේ මූලධර්මය වන්නේ වායු සහ රුධිර ප්රවාහ අතර CO 2 සහ O 2 හුවමාරු කිරීම, එකිනෙකට ප්රතිවිරුද්ධව යොමු කිරීමයි.

සහල්. 12. මාළු (I) සහ භූමිෂ්ඨ පෘෂ්ඨවංශීන්ගේ (II) රුධිර සංසරණ යෝජනා ක්රමය:
1 - ශාඛා ධමනි; 2 - කැරොටයිඩ් ධමනිය; 3 - පෘෂ්ඨීය aorta; 4 - උදර aorta; 5 - පුඵ්ඵුසීය ධමනිය; 6 - Cuvier duct; 7 - ඉදිරිපස කාර්දිනල් (ජුගුලර්) නහර; 8 - පසුපස කාර්දිනල් නහර; 9 - පසුපස vena cava; 10 - අක්මාව නහර; 11 - පෙනහළු නහර; 12 - අක්මාවේ ද්වාර නහර; 13 - උප බඩවැලේ නහර
සංසරණ අවයව. පෘෂ්ඨවංශීන්ගේ රුධිර සංසරණ පද්ධතිය, හිස්කබල නොවන අයගේ මෙන්, වසා ඇත. සංසරණ පද්ධතිය පාර්ශ්වීය තහඩු වල අභ්යන්තර තහඩු වලින් තැන්පත් කර ඇත (ලැන්ස්ලට් සංවර්ධනය බලන්න). එය අන්තර් සම්බන්ධිත රුධිර වාහිනී වලින් සමන්විත වන අතර, රළු යෝජනා ක්රමයකින් ටන්ක දෙකකට අඩු කළ හැකිය: පෘෂ්ඨීයඑහිදී රුධිරය හිස සිට වලිගය දක්වා ගලා යයි, සහ උදරීයදිගේ එය ප්රතිවිරුද්ධ දිශාවට ගමන් කරයි. හිස්කබල නොවන (රූපය 12) මෙන් නොව, පෘෂ්ඨවංශීන් තුළ, රුධිරයේ චලනය හෘදයේ ක්රියාකාරිත්වය සමඟ සම්බන්ධ වේ.
හදවත ඝන බිත්ති සහිත මාංශ පේශි බෑගයක්, කොටස් කිහිපයකට බෙදා ඇත - කුටි. හදවතේ ප්රධාන කොටස් වන්නේ රුධිරය ලබා ගන්නා කර්ණිකාව සහ ශරීරය හරහා එය මෙහෙයවන කශේරුකාවයි. පෘෂ්ඨවංශීන්ගේ විවිධ පන්තිවල හෘද කුටි ගණන වෙනස් වේ.
කලලරූපීව, හදවත පැන නගින්නේ උදරීය aorta හි පසුපස කොටසෙහි ප්රසාරණයක් ලෙස වන අතර එය මෙම ස්ථානයේ වක්ර ලූපයකට දඟර ගනී. ලූපයේ ඉදිරිපස කොටස මතු වේ කශේරුකාවහදවත්, පිටුපස - ආලින්දය.
හෘදයේ ස්වයංක්රීය මාදිලියේ ක්රියා කරන ඉරි සහිත මාංශ පේශි ඇති අතර එහි හැකිලීම ස්වේච්ඡා ආවේගයන්ට යටත් නොවේ. හෘදයේ මානයන් එහි කාර්යයේ තීව්රතාවයට සම්බන්ධ වන අතර, පෘෂ්ඨවංශීන්ගේ ශ්රේණියේ ශරීරයේ මානයන්ට සාපේක්ෂව එහි මානයන් වැඩි වේ (වගුව 1).
වගුව 1
විවිධ පංතිවල පෘෂ්ඨවංශීන්ගේ හෘද දර්ශකය
රුධිර වාහිනී පද්ධති දෙකකට බෙදා ඇත: ධමනිහෘදයෙන් රුධිරය ගලා යන, සහ ශිරාඑමගින් රුධිරය හදවතට නැවත පැමිණේ. පෘෂ්ඨවංශීන්ගේ සංකූලතාවයේ ක්රියාවලියේදී, රුධිර සංසරණයේ එක් චක්රයක් ඇති සතුන්ගේ සිට රුධිර සංසරණ කව දෙකක අයිතිකරුවන් දක්වා සංක්රමණය වේ.
එහි ස්වභාවය අනුව, රුධිරය අන්තර් සෛලීය අවකාශයෙන් රුධිරයට විනිවිද යන සම්බන්ධක පටක වේ.
පෘෂ්ඨවංශීන්ගේ රුධිරය අවර්ණ ද්රවයකින් සමන්විත වේ - ප්ලාස්මා, එහි පිහිටුවා ඇති රුධිරයේ මූලද්රව්ය පිහිටා ඇත: රතු රුධිර සෛල හෝ එරිත්රෝසයිට්, ඩයි අඩංගු - හිමොග්ලොබින් සහ සුදු රුධිර සෛල - ලියුකෝසයිට්. රතු රුධිර සෛල ඔක්සිජන් රැගෙන යන බැවින් රුධිර ඔක්සිකරණය සමඟ සම්බන්ධ වේ. ශරීරයට ඇතුල් වී ඇති ක්ෂුද්ර ජීවීන් විනාශ කිරීම සඳහා ලේයිකොසයිට් සම්බන්ධ වේ. මීට අමතරව, රුධිරයේ පට්ටිකා ඇති අතර, රුධිර කැටි ගැසීමේ දී වැදගත් කාර්යභාරයක් ඉටු කරයි, මෙන්ම අනෙකුත් සෛල. පෘෂ්ඨවංශීන්ගේ පරිණාමීය මාලාවේ රුධිර ස්කන්ධය වැඩි වේ (වගුව 2).
වගුව 2
විවිධ පංතිවල පෘෂ්ඨවංශීන්ගේ ශරීර ස්කන්ධයට සාපේක්ෂ රුධිර ස්කන්ධය
(ප්රොසර් සහ බ්රවුන්ට පසුව, 1967; ප්රොසර්ට පසුව, 1978)
සංසරණ පද්ධතිය බහුකාර්ය වේ. අවයව, පටක, ඔක්සිජන් සෛල, කාබනික සහ ඛනිජ ද්රව්ය, ද්රව සහ දිරාපත්වන නිෂ්පාදන ඉවත් කිරීම, විෂ ද්රව්ය, කාබන් ඩයොක්සයිඩ්, අන්තරාසර්ග ග්රන්ථි වල හෝමෝන මාරු කිරීමේදී එය ලැබීමට සම්බන්ධ වේ.
සංසරණ පද්ධතිය සමඟ, පෘෂ්ඨවංශීන්ට ඒ හා සම්බන්ධ තවත් සනාල පද්ධතියක් ඇත - වසා ගැටිති. එය සමන්විත වේ වසා වාහිනී සහ වසා ග්රන්ථි. වසා පද්ධතිය වසා නැත. එහි විශාල යාත්රා පමණක් ස්වාධීන බිත්ති ඇති අතර, ඒවායේ ශාඛා විවිධ අවයවවල අන්තර් සෛල අවකාශයට විවෘත වේ. වසා ගැටිති වල අවර්ණ තරලයක් අඩංගු වේ - වසා ගැටිති, වසා ග්රන්ථි වල පිහිටුවා ඇති ලිම්ෆොසයිට් පාවෙන. වසා ගැටිති වල චලනය තීරණය වන්නේ බිත්තිවල හැකිලීම, විශාල නාල වල සමහර කොටස් (ඊනියා වසා හෘදය) සහ විවිධ අවයවවල භාජන මත වරින් වර වෙනස් වන පීඩනය මගිනි.
වසා පද්ධතිය රුධිරය හා පටක අතර ද්රව්ය හුවමාරු කිරීමේ අතරමැදියෙකු ලෙස සේවය කරයි.
ස්නායු පද්ධතිය. ස්නායු පද්ධතියේ කාර්යයන් වන්නේ බාහිර උත්තේජක සංජානනය සහ සෛල, අවයව, පටක වලට නැගී එන උද්දීපනය සම්ප්රේෂණය කිරීම මෙන්ම තනි ඉන්ද්රිය පද්ධති සහ සමස්තයක් ලෙස ශරීරයේ ක්රියාකාරකම් ඒකාබද්ධ කිරීම හා සම්බන්ධීකරණය කිරීම තනි ක්රියාකාරී ජීවන රටාවකට ය. පද්ධතියක්. කළල වශයෙන්, පෘෂ්ඨවංශීන්ගේ ස්නායු පද්ධතිය පැන නගින්නේ, හිස්කබල නොවන ඒවායේ මෙන්, කලලයේ පෘෂ්ඨීය පැත්තේ ඇති ectoderm හි ඇති හිස් නලයක ස්වරූපයෙන් (රූපය 13). පසුව, එහි අවකලනය සිදු වේ, එය ගොඩනැගීමට තුඩු දෙයි: a) මධ්යම ස්නායු පද්ධතිය, මොළය සහ සුෂුම්නාව මගින් නිරූපණය කෙරේ, b) පර්යන්ත ස්නායු පද්ධතිය, මොළයේ සහ සුෂුම්නාවෙන් විහිදෙන ස්නායු වලින් සමන්විත වන අතර c) සානුකම්පිත ස්නායු පද්ධතිය, කොඳු ඇට පෙළ අසල පිහිටා ඇති ස්නායු නෝඩ් පදනමින් සමන්විත වන අතර කල්පවත්නා කෙඳි මගින් සම්බන්ධ වේ.
මොළය පෘෂ්ඨවංශීන් තුළ කොටස් පහකින් නිරූපණය කෙරේ: ඉදිරිපස, අතරමැදි, මැද, මස්තිෂ්ක සහ දිගටිමොළය. එය ඉක්මනින් ප්රාථමික මස්තිෂ්ක වෙසිලි තුනකට බෙදෙන ස්නායු නාලයේ ඉදිරිපස කොටස ඉදිමීමේ ස්වරූපයෙන් කලලයක් තැන්පත් කර ඇත (රූපය 14). අනාගතයේ දී, පළමු මස්තිෂ්ක මුත්රාශයේ ඉදිරිපස ඉදිරිපස ඇති කරයි; එහි පිටුපස කොටස diencephalon බවට පරිවර්තනය වේ. මැද මොළය සෑදී ඇත්තේ දෙවන මස්තිෂ්ක මුත්රාශයේ සිටය. නෙරා යාමෙන්

සහල්. 13. පෘෂ්ඨවංශීන්ගේ මධ්යම ස්නායු පද්ධතියේ වර්ධනයේ අනුප්රාප්තික අවධීන් (ක්රමානුකූල හරස්කඩ):
ectoderm හි I-II අවකලනය; ස්නායු තහඩුවේ III ආක්රමණ, IV-V - ස්නායු නාලය හුදකලා කිරීම, 1 epidermis; 2 ganglion තහඩුව

සහල්. 14. මොළයේ කොටස් වර්ධනය (යෝජනා ක්රමය):
I - බුබුලු තුනක අදියර (ඇස් බුබුලු සහිත), II - දෙපාර්තමේන්තු පහක අදියර (ඇස් කෝප්ප සහිත), 1 - forebrain, 2 - diencephalon; 3 - මැද මොළය; 4 - මස්තිෂ්ක; 5 - medulla oblongata; 6 - අක්ෂි කුසලාන; 7 - අක්ෂි බුබුල
තුන්වන මස්තිෂ්ක මුත්රාශයේ වහලය මස්තිෂ්ක සාදයි, ඒ යටතේ medulla oblongata පිහිටා ඇත. ඉදිරි මොළය වම් සහ දකුණු අර්ධ වශයෙන් තවදුරටත් බෙදී ඇත.
ස්නායු නාලයේ හිස කොටසෙහි වර්ධනය හා අවකලනය සමග සමගාමීව, neurocoel හි අනුරූප පරිවර්තනයක් සිදු වේ. පූර්ව මොළයේ අර්ධගෝලයේ එහි දිගු දෙකක් ලෙස හැඳින්වේ පාර්ශ්වීය කශේරුකාමොළය. මොළයේ අතරමැදි කොටසෙහි නියුරොකොයෙල්හි පුළුල් වූ කොටස ලෙස නම් කර ඇත තුන්වන කශේරුකාව, මැද මොළය කුහරය - ලෙස සිල්වියන් ජලධරය, medulla oblongata හි කුහරය - ලෙස හතරවන කශේරුකාව, හෝ rhomboid fossa(රූපය 14). යුගල 10ක් හෝ 12ක් මොළයෙන් පිටවේ කපාල ස්නායු.
පෙර මොළේ ඉදිරිපස සමමිතිකව පිහිටා ඇති නෙරා යාමේ දෙකක් ඇත, එයින් පළමු හිස ස්නායු යුගලය පිටත් වේ - සුවඳ විලවුන්(මම). Diencephalon පතුලේ සිට පිටත් වේ දෘශ්යස්නායු (හිස ස්නායු දෙවන යුගලය, II).
ඩයන්ස්ෆලෝන්හි වහලය මත, කකුල් මත වාඩි වී සිටින නෙරා යාමේ දෙකක් වර්ධනය වේ: ඉදිරිපස - ප්රාචීර ඉන්ද්රියසහ ආපසු - epiphysis.
යුගල නොකළ නෙරා යාමක් ඩයන්ස්ෆලෝන් පතුලේ සිට වර්ධනය වේ - පුනීලය, එය ව්යුහයෙන් සහ ක්රියාකාරීත්වයෙන් සංකීර්ණ වූ සැකැස්මකට යාබදව - පිටියුටරි. ඉදිරිපස පිටියුටරි ග්රන්ථිය වර්ධනය වන්නේ මුඛ කුහරයේ එපිටිලියම් වලින්, පසුපස - මෙඩුල්ලා වලින්. එය එහි පිහිටා ඇත හයිපොතලමස්.
මැද මොළයේ වහලය යුගල ඉදිමීම් සාදයි - දෘශ්ය පෙති (tubercles). මොළයේ ස්නායු වල තුන්වන යුගලය මැද මොළයෙන් පිටත් වේ ( අක්ෂි මෝටරය, III). සිව්වන හිස ස්නායු යුගල ( අවහිර කරන්න, IV) මැද සහ medulla oblongata අතර මායිමෙන් පිටත් වේ, අනෙකුත් සියලුම හිස ස්නායු (V - X - XII) medulla oblongata වෙතින් පිටත් වේ.
සුෂුම්නාව medulla oblongata වලින් තියුණු ලෙස සීමා නොකෙරේ. කොඳු ඇට පෙළේ මධ්යයේ (ඉන්ද්රියයේ ප්රධාන අක්ෂය දිගේ), නියුරෝකොයෙල් සංරක්ෂණය කර ඇති අතර එය පෘෂ්ඨවංශීන් තුළ නමින් හැඳින්වේ. කොඳු ඇට පෙළ.
කොඳු ඇට පෙළේ ස්නායු කොඳු ඇට පෙළෙන් පාරමිතිකව (කොටස් ගණන අනුව) පිටත් වේ. ඒවා මූල දෙකකින් ආරම්භ වේ: පෘෂ්ඨීය - සංවේදී සහ උදර - මෝටර්. මෙම මූලයන් සුෂුම්නාවෙන් පිටවී ටික වේලාවකට පසු විලයනය වී කොඳු ඇට පෙළේ ස්නායු සාදයි, පසුව එය නැවත පෘෂ්ඨීය සහ උදර අතු වලට බෙදී යයි.
ඉන්ද්රියයන්. මෙම අවයව සමූහය කලලරූපයේ විවිධ කොටස්වල ව්යුත්පන්නයන් ලෙස සහ එහි වර්ධනයේ විවිධ අවස්ථා වලදී පැන නගී. මේවා සුවඳ, දර්ශනය, ශ්රවණය, වෙස්ටිබුලර් උපකරණ, පාර්ශ්වීය රේඛාවේ අවයව, රසය, ස්පර්ශය, පෘථිවි චුම්බක ක්ෂේත්රය වටහා ගන්නා විශේෂිත අවයව, විද්යුත් ක්ෂේත්ර, තාප විකිරණ යනාදිය වේ.
ආඝ්රාණ අවයව. සුවඳ දැනීම මොළයේ පැරණිතම කාර්යයක් බව විශ්වාස කෙරේ. ගන්ධයේ අවයව ස්නායු තහඩුව සමඟ සමගාමීව ectoderm ඝණ වීම ලෙස තබා ඇත. සමාන්තරව, මොළයේ හිස්කබලේ කොටසක් වන සුවඳ කැප්සියුලවල ඇටසැකිල්ල සෑදී ඇත. මුලදී, සුවඳ කැප්සියුල බාහිර පරිසරය සමඟ පමණක් සන්නිවේදනය කරන අතර බාහිර නාස්පුඩු ඇත. පසුව, භෞමික පැවැත්ම නිසා, නාස්පුඩු හරහා බවට පත් වේ.
දර්ශනයේ අවයවපැරණි සංවේදී ඉන්ද්රියයන්ටද අයත් වේ. ආලෝක සංවේදී පිළිගැනීමක් chordates පරිණාමයේ ඉතා මුල් අවධියේදී සිදු වේ (lancelet මතක තබා ගන්න) සහ මුල් කළල උත්පාදනය තුළ පිහිටුවා ඇත. පෘෂ්ඨවංශීන්ගේ දර්ශනයේ අවයව යුගල සහ යුගල නොකළ ලෙස බෙදා ඇත. දෙකම ඩයන්ස්ෆලෝන්හි වර්ධනය වේ. යුගල කරන ලද ඇස් ඩීන්ස්ෆලෝන්හි පාර්ශ්වීය කොටස්වල වර්ධනයක් ලෙස තබා ඇත, යුගල නොකළ - අනුක්රමිකව ඩයින්ස්ෆලෝන්හි වහලයේ (පයිනල් ග්රන්ථිය සහ ප්රාචීර ඉන්ද්රිය) පිහිටා ඇත. යුගල ඇස් තැබීම මොළයේ හිස්කබලේ කොටසක් වන ඔවුන් වටා දෘශ්ය කැප්සියුල සෑදීම සමඟ ඇත (රූපය 15, 16).
ශ්රවණ අවයවපෘෂ්ඨවංශීන් තුළ සංකීර්ණ සම්භවයක් ඇත. පරිණාමයේ මුල්ම අභ්යන්තර කණ වේ

සහල්. 15. අක්ෂි වර්ධනයේ අනුප්රාප්තික අවධීන්:
1 - diencephalon; 2 - කාචය සාදන එපීඩර්මිස්; 3 - අක්ෂි කෝප්පය සෑදීමේ ආරම්භය; 4 - අක්ෂි කෝප්පයේ කකුල; 5 - කාච; 6 - කෝනියා; 7 - දෘෂ්ටි විතානය; 8 - වර්ණක ෂෙල්; 9 - choroid සහ sclera සෑදෙන මෙසෝඩර්ම් සෛල

සහල්. 16. මිනිස් ඇසේ සජිටල් කොටස:
1 - කෝනියා; 2 - අයිරිස්; 3 - ඉදිරිපස කුටිය; 4 - ඇසේ චලනය වන මාංශ පේශිවලින් එකක්; 5 - වීදුරු ශරීරය; 6 - දෘෂ්ටි ස්නායුව; 7 - ස්ක්ලෙරා; 8 - choroid; 9 - දෘෂ්ටි විතානය; 10 - සිලියරි මාංශ පේශි; 11 - සිලියරි ශරීරය
කලලයේ ectoderm තුළ තැන්පත් කර ඇති අතර, කුහරයක ස්වරූපයෙන් ගැඹුරු වන අතර හැඩය ගනී auditory vesicleශ්රවණ කැප්සියුලයේ වැතිර සිටී. ශ්රවණ වෙසිලය සංකෝචනය වීමෙන් කොටස් දෙකකට බෙදී ඇත. ඉහළ කොටස වෙස්ටිබුලර් උපකරණය බවට පත්වේ. එය සමතුලිතතාවයේ ඉන්ද්රියයි. පෘථිවියේ ත්රිමාණ අවකාශයේ ශරීරයේ පිහිටීම දැනීමට එය ඔබට ඉඩ සලසයි. මෙම ශරීරය - 3 අර්ධ වෘත්තාකාර ඇලඅභ්යන්තර කණ තුළ (රූපය 17, 18). ශ්රවණ වෙසිලයේ පහළ කොටස අභ්යන්තර කණ වේ - ශ්රවණ මල්ල.
ගොඩබිම ආශ්රිතව පෘෂ්ඨවංශීන් මතුවීමේ ප්රමාද අවධියේදී මැද සහ පිටත කණ සෑදී ඇත.
පාර්ශ්වීය රේඛා අවයවප්රාථමික ජලජ පෘෂ්ඨවංශීන්ගේ පමණක් ලක්ෂණය, ectoderm හි ද තැන්පත් කර ඇත. ඒවා දිග හැරෙන කට්ට

සහල්. 17. අභ්යන්තර කණ වර්ධනයේ අනුප්රාප්තික අවධීන්:
I - auditory placode, II - fossa, III සහ IV - කොටසෙහි වෙසිලිය; V සහ VI - අර්ධ වෘත්තාකාර ඇල සෑදීම; 1 - ගොළුබෙල්ලෙකුගේ මූලාරම්භය; 2 - අර්ධ වෘත්තාකාර ඇල; 3 - එන්ඩොලිම්ෆැටික් නාලය; 4 - රවුම් බෑගය; 5 - ඕවලාකාර මල්ලක්
හිස දෙපස සහ ශරීරය දිගේ. මෙම කට්ට අස්ථි කොරපොතු වලින් ආවරණය වී හෝ නොතිබිය හැකිය. පාර්ශ්වික රේඛීය අවයව කම්පන ප්රභවයන් අසල ආලෝක චලනයන් සහ ජල කම්පන වටහා ගනී: ධාරා වල වේගය සහ දිශාව, තමන්ගේම ශරීරයේ චලනයන් සහ ජලයේ සත්වයාගේ මාර්ගයේ වස්තූන් තිබීම. ඒවා භූ කම්පන සංවේදක පද්ධතියකි.
රස අවයවඑන්ඩොඩර්ම් වල තැන්පත් කර ඇති අතර පරාසය තුළ පරිභෝජනය කරන ආහාරවල රසය වටහා ගනී: පැණිරස, කටුක, ඇඹුල්, ලුණු. ඒවා මුඛ කුහරය තුළ රස අංකුර මත පිහිටා ඇත.
ඉන්ද්රියයන්. ඒවාට සංවේදී සෛල නොමැත, නමුත් සමෙහි අතු බෙදී ඇති ස්නායු අවසානය වන අතර ඒවා ස්පර්ශයෙන් පාරිසරික වස්තූන් වටහා ගනී.

සහල්. 18. පටල සහිත ශ්රවණ ලිහිණියේ යෝජනා ක්රමය:
1, 2, 3 - ඉදිරිපස, බාහිර සහ පසුපස අර්ධ වෘත්තාකාර ඇල, 4 - එන්ඩොලිම්ෆැටික් නාලිකාව, 5 - ශ්රවණ ලප, 6 - රවුම් මල්ල, 7 ඕවලාකාර මල්ල, 8 - ඔටොලිත්

සහල්. 19. සනාල පටලැවිල්ල සහිත බෝමන්ගේ කැප්සියුලය:
1 - afferent රුධිර නාල, 2 - පිටාර රුධිර නාල, 3 - Bowman's capsule, 4 - සනාල පටලැවිල්ල, 5 - ප්රාථමික මුත්රා, 6 - වකුගඩු නාල, 7 - වකුගඩු නාලයේ (nephrostome) පුනීල ශරීර කුහරයට විවෘත වේ.

සහල්. 20. පෘෂ්ඨවංශීන්ගේ මුත්රා පද්ධතියේ යෝජනා ක්රමය:
I (පිරිමි) සහ II (ගැහැණු) - මෝරුන් සහ උභයජීවීන්, III (පිරිමි) සහ IV (ගැහැණු) - උරගයින් සහ පක්ෂීන්, V (පිරිමි) සහ VI (ගැහැණු) - ක්ෂීරපායින්, 1 - ප්රොනෙෆ්රොස් (ප්රොනෙෆ්රොස්); 2 - වෘෂණ ; 3 - vas deferens, 4 - mesonephros (ප්රාථමික වකුගඩු), 5 - hindgut, 6 - මුත්රාශය, 7 - cloaca, 8 - පුනීල, 9 - ඩිම්බකෝෂය, 10 - Muller ඇල, 11 - epididymis (මෙසෝවේ ඉදිරිපස කොටසෙහි ඉතිරි කොටස - nephros), 12 - vas deferens, 13 - metanephros (ද්විතියික වකුගඩු), 14 ද්විතීයික මුත්රාශය, 15 mesonephros rudiment, 16 oviduct, 17 ඩිම්බ කෝෂ, 18 ඩිම්බ කෝෂ බිත්තියේ ග්රන්ථි මගින් ස්රාවය වන ප්රෝටීන් 18, ගර්භාෂ 19 - embr, 20 ගර්භාෂය 21 - යෝනි මාර්ගය, 22 - ලිංගික සයිනස්, 23 - පුරස්ථි ග්රන්ථි; 24 - ශුක්ර තරලය, 25 - vas deferens, 26 - epididymis, 27 - perineum; 28 - Muller ඇලෙහි rudiment, 29 - mesonephros හි ඇල (ප්රාථමික වකුගඩු); 30 - copulatory අවයව (ශිෂේණය), 31 - ගුදය
බැහැර කිරීමේ අවයව. සියලුම පෘෂ්ඨවංශීන්ට බැහැර කිරීමේ අවයව ඇත වකුගඩුශරීරයෙන් යූරියා හෝ යූරික් අම්ලය ආකාරයෙන් අතිරික්ත ජලය, ඛනිජ ලවණ සහ නයිට්රජන් පරිවෘත්තීය වියෝජන නිෂ්පාදන ඉවත් කිරීමට නිර්මාණය කර ඇත. ඒවා මෙසෝඩර්මල් සම්භවයක් ඇති අතර ඒවා සෝමයිට් වල පිටත බිත්තිය මත තබා ඇත. කෙසේ වෙතත්, පෘෂ්ඨවංශීන්ගේ විවිධ කණ්ඩායම්වල වකුගඩු වල ක්රියාකාරිත්වයේ ව්යුහය සහ යාන්ත්රණය සමාන නොවේ. පෘෂ්ඨවංශීන්ගේ පරිණාමය ක්රියාවලියේදී, වකුගඩු වර්ග තුනක් වෙනස් වේ: හිස, හෝ pronephros (pronephros), කඳ, හෝ ප්රාථමික, වකුගඩු (මෙසොනෙෆ්රොස්) හා ශ්රෝණිය, හෝ ද්විතියික, වකුගඩු (metanephros) විවිධ වර්ගයේ වකුගඩු වල බැහැර කිරීමේ ක්රියාවලියේ විවිධ මූලධර්ම ඇත: ශරීර කුහරයෙන් බැහැර කිරීම, මිශ්ර පිටකිරීම් (ශරීර කුහරයෙන් සහ රුධිරයෙන්) සහ අවසාන වශයෙන්, රුධිරයෙන් පමණි. ඒ සමගම, ජලය නැවත අවශෝෂණය කිරීමේ යාන්ත්රණයේ වෙනස්කම් සිදු වේ. ශරීර කුහරයෙන් ජලය මුදා හැරීම සහ එහි දිය වී ඇති ප්රෝටීන් පරිවෘත්තීය නිෂ්පාදන බොහෝ පුනීල හැඩැති නෙෆ්රොස්ටෝමා ශරීර කුහරයට විවර වීම නිසා සිදුවිය හැකිය. රුධිරයෙන් බැහැර කිරීම වකුගඩු වල Malpighian සිරුරු හරහා සිදු වේ. වකුගඩු වලින් බැහැර කරන නාලිකා ලෙස හැඳින්වේ වෘක නාලිකා, ඒවා ප්රතිස්ථාපනය වේ ureters. බොහෝ පෘෂ්ඨවංශීන් සතුව ඇත මුත්රාශය. ප්රාථමික ජලජ වලදී, ඇමෝනියා ගිලිනු හරහා බැහැර කළ හැක.
ලිංගික අවයව. පෘෂ්ඨවංශීන්ගේ ලිංගික ග්රන්ථි - කාන්තාවන්ගේ ඩිම්බ කෝෂ සහ පිරිමින්ගේ වෘෂණ - සාමාන්යයෙන් යුගලනය වේ. මෙම මූලද්රව්යය සොමයිට් සහ පාර්ශ්වීය තහඩුවට බෙදීමේ ස්ථානයේ ඇති මෙසෝඩර්ම් කොටසේ සිට ඒවා වර්ධනය වේ.
මුලදී (හකු රහිතව) ලිංගික ග්රන්ථිවලට බැහැර කරන නාලිකා නොතිබූ අතර ප්රජනක නිෂ්පාදන ලිංගික ග්රන්ථි වල බිත්තිවල සිදුරු හරහා ශරීර කුහරයට වැටී ඇති අතර එහිදී ඒවා විශේෂ සිදුරු හරහා බාහිර පරිසරයට බැහැර කරයි. පසුව, ලිංගික පත්රිකාව මතු වූ අතර එය පිරිමින් තුළ බැහැර කිරීමේ අවයව (වුල්ෆ්ගේ ඇල) සමඟ සම්බන්ධ වේ. තවද කාන්තාවන් තුළ එය ඩිම්බකෝෂයක් ලෙස ක්රියා කරයි මුලර් නාලිකාව, බාහිර පරිසරය සමඟ සමස්ථයේ සම්බන්ධය පවත්වාගෙන යන.
පෘෂ්ඨවංශීන්- lat. පෘෂ්ඨවංශීන්, සත්ව ලෝකයේ මෙම නියෝජිතයින්ගේ ලක්ෂණයක් වන්නේ සැබෑ අස්ථි හෝ කාටිලේජීය ඇටසැකිල්ලක් තිබීමයි, එහි පදනම කොඳු ඇට පෙළයි. මෙම කණ්ඩායම ඉතා විවිධාකාර වන අතර පහත සඳහන් සතුන් වර්ග ඇතුළත් වේ: මාළු, උභයජීවීන්, උරගයින්, කුරුල්ලන් සහ ක්ෂීරපායින්.
පෘෂ්ඨවංශීන්ගේ සම්භවය සහ සත්ව විද්යාව
ඉහළ chordates දී - පෘෂ්ඨවංශීන්, හෝ හිස් කබල, notochord ක්රමයෙන් කශේරුකා මගින් ප්රතිස්ථාපනය වේ. ඔවුන්ගේ ඇටසැකිල්ලේ පදනම වන්නේ කොඳු ඇට පෙළයි. පෘෂ්ඨවංශීන්ට හිස් කබලක් ඇත. ඔවුන්ගේ සංසරණ පද්ධතියට හදවත ඇතුළත් වේ. චෝඩේට් වල පහළ කණ්ඩායම් මෙන් නොව, පෘෂ්ඨවංශීන් සක්රියව ආහාර සොයමින් අල්ලා ගනී. එවැනි ක්රියාකාරී පෝෂණය පැවැත්ම සඳහා අරගලයේ දී පෘෂ්ඨවංශීන්ට සැලකිය යුතු වාසි ලබා දුන්නේය.
පෘෂ්ඨවංශීන් පරිණාමය වූයේ ප්රාථමික නොවන හිස්කබල ලෙස හැඳින්වෙන ලැන්ස්ලට් වලට සමාන ප්රාථමික chordates වලින්.
මාළු සම්භවය
මීට වසර මිලියන 400 කට පමණ පෙර මාළු පන්තියේ නියෝජිතයන් මතු විය. අප අධ්යයනය කරන සතුන් අතරින් වඩාත් දුර්වල ලෙස සංවිධානය වූ පෘෂ්ඨවංශීන් මේවාය. ඒ අතරම, දිගු පරිණාමයකදී, මාළු ජලජ පරිසරයේ ජීවිතයට පරිපූර්ණ ලෙස අනුවර්තනය වී ඇත. ක්රියාකාරී චලනයන්, මාංශ පේශී, අභ්යන්තර ඇටසැකිල්ල, ගිල් උපකරණ, මොළය සහ සංවේදී ඉන්ද්රියන්ගේ අවයව විශේෂ වර්ධනයකට පැමිණ ඇත.
මසුන්ගේ පරිණාමයේ අත්යවශ්ය මොහොතක් වූයේ යුගල නොකළ, යුගල කළ වරල් සෑදීමයි. ඉහළ පෘෂ්ඨවංශීන්ගේ පසුකාලීන පරිණාමය තුළ මෙය අතිශයින් වැදගත් කාර්යභාරයක් ඉටු කළේය.
පෘෂ්ඨවංශික පෘෂ්ඨවංශීන්ගේ පෙනුමට හේතු වූ සත්ව සංවර්ධනයේ ශාඛාව ද පැරණි මසුන්ගෙන් ඉවත් විය.
උභයජීවීන්ගේ සම්භවය
භූමිෂ්ඨ පෘෂ්ඨවංශීන්ගේ පෙනුමට බොහෝ කලකට පෙර, පෘථිවිය පුරාණ මත්ස්යයන් - lobe-finned මාළු විසින් වාසය කරන ලදී. මෙම lobe-finned මසුන්ගේ එක් ශාඛාවක්, භෞමික පැවැත්මට අනුවර්තනය වීම, ප්රාථමික උභයජීවීන් බිහි විය. ගොඩබිම ජීවිතයට සංක්රමණය වීමත් සමඟ, එවැනි ක්රොසොප්ටෙරිජියන්ට ඔවුන්ගේ ගිලන් නැති වී පෙනහළු වලින් හුස්ම ගැනීමට පටන් ගත් අතර, වරල් වෙනුවට ඔවුන් ඇඟිලි පහේ අත් පා සෑදූ අතර ඔවුන්ගේ සංසරණ පද්ධතිය වඩාත් සංකීර්ණ විය.
සත්ව පොසිල අධ්යයනයෙන් සනාථ වන පරිදි, පළමු උභයජීවීන්, lobe-finned මාළු වලින් පැවත එන අතර, පෘථිවි ඉතිහාසයේ පුරාණ යුගයේ එක් කාල පරිච්ඡේදයක් අවසානයේ පෘථිවියේ දර්ශනය විය - ඩෙවෝනියානු යුගය. මීළඟ කාලපරිච්ඡේදයේ දී, කාබන්ඩයොක්සයිඩ්, කාලපරිච්ඡේදය, උභයජීවීන් දැනටමත් විශේෂවල පොහොසත්කම සහ විවිධත්වය මගින් කැපී පෙනෙන අතර විශාල ව්යාප්තියකට ළඟා විය.
භෞමික වෘක්ෂලතාදිය (ගස් වැනි පර්ණාංග, අශ්වාරෝහක සහ ක්ලබ් පාසි) සුවිශේෂී ලෙස පොහොසත් වර්ධනයක් මගින් කැපී පෙනෙන කාබොනිෆරස් යුගයේ උණුසුම් හා තෙතමනය සහිත දේශගුණය උභයජීවීන්ගේ වර්ධනයට අනුග්රහය දැක්වීය. කෙසේ වෙතත්, එවකට පෘථිවියේ ඉහළ පෘෂ්ඨවංශීන් (පක්ෂීන්, ක්ෂීරපායින්) සිටියේ නැත.
පුරාණ ෆොසිල උභයජීවීන් අයත් වූයේ වඳ වී ගිය ස්ටෙගෝසෙෆල් කාණ්ඩයට වන අතර එහි අර්ථය "ෂෙල්-හිස්" යන්නයි. මෙම සතුන්ට මෙම නම ලැබුණේ ඔවුන්ගේ හිස් කබල පිටතින් ඝන කවචයකින් ආවරණය වී ඇති බැවිනි. කවචය ඔවුන්ගේ ශරීරය අර්ධ වශයෙන් ආවරණය කළේය.
ස්ටෙගෝසෙෆාලියන් ඔවුන්ගේ සංවිධානයේ රඳවා තබා ඇති මසුන්ගේ බොහෝ සලකුණු ඔවුන්ගේ සම්භවය සෘජුවම තහවුරු කරයි.
මාළු වලින් උභයජීවීන්ගේ සම්භවය පිළිබඳ නොඅඩු ඒත්තු ගැන්විය හැකි සාක්ෂි වන්නේ නූතන උභයජීවීන්ගේ කීටයන්ගේ මසුන් සමඟ ඇති සමානකම් - ටැඩ්පෝල්ස්: මාළු වැනි ශරීර හැඩය, පෞච්ඡ වරල් තිබීම, ගිල් හුස්ම ගැනීම යනාදිය.
උරගයන්ගේ සම්භවය
නූතන උභයජීවීන් හෝ උභයජීවීන් සමඟ, ස්ටෙගෝසෙෆාලියන් උරගයින් සඳහා පදනම දැමීය. උරගයින්ගේ මතුවීම පුරාණ යුගයේ අවසානයේ ඇති වූ දේශගුණික විපර්යාස සමඟ සමීපව සම්බන්ධ වේ. මෙම අවස්ථාවේදී පෘථිවියේ දේශගුණය වියළි වේ, ජල කඳන් වියළී යයි, තෙත් බිම් විශාල භූමි ප්රදේශ බවට පත්වේ.
මෙය පුරාණ උභයජීවීන්ගේ ක්රමානුකූල වෙනසක් සහ ප්රාථමික උරගයින් ඔවුන්ගෙන් මතු වීමට හේතු විය.
උරග පන්තිය සැබෑ භූමිෂ්ඨ පෘෂ්ඨවංශීන්ගේ පළමු පන්තියයි.
වේගයෙන් වර්ධනය වෙමින්, උරගයින් ඉක්මනින්ම ප්රමුඛ ස්ථානයක් ගත් අතර උභයජීවීන් පසුබිමට තල්ලු කළේය.
උරගයින්ගේ විශේෂයෙන් වේගවත් සංවර්ධනයක් ඊළඟ - මැද යුගයේ සිදු විය. පෙර ද්රව්ය වලින් අප දැනටමත් දන්නා පරිදි, උරගයින් අතිවිශාල ප්රමාණයකින් යුක්ත වූ අතර විවිධ විශේෂයන් විසින් නියෝජනය කරන ලද්දේ මේ අවස්ථාවේදීය. ඒවා ගොඩබිම, ජලය සහ වාතයේ බහුලව දක්නට ලැබිණි.
පොසිල උරගයින් සුවිශේෂී උනන්දුවක් දක්වයි. පසුව, සත්ව ලෝකයේ එම ශාඛා පැරණි උරගයන්ගෙන් මතු වූ අතර, නවීන උරගයින්ට අමතරව, කුරුල්ලන් සහ ක්ෂීරපායින් ද ආරම්භ විය.
කුරුල්ලන්ගේ සම්භවය
නූතන පක්ෂීන්ගේ පුරාණ මුතුන් මිත්තන් වන Archeopteryx (පළමු තටුව" ලෙස පරිවර්තනය කර ඇත), එහි ව්යුහය සමඟ අප දැනටමත් හුරුපුරුදු වී ඇති අතර, සමහර උරගයින් පියාසර කිරීමට අනුවර්තනය වීමේ ප්රති result ලයක් ලෙස පැන නගී.
Archeopteryx සොයා ගැනීමෙන් කුරුල්ලන් උරගයන්ගෙන් පැවත එන බව ඒත්තු ගැන්විය හැකිය. උරගයින් සමඟ කුරුල්ලන්ගේ සම්බන්ධතාවය ඔවුන්ගේ ව්යුහය හා සංවර්ධනයේ බොහෝ පොදු ලක්ෂණ මගින් ද සාක්ෂි දරයි.
Archeopteryx හි ව්යුහය අධ්යයනයෙන් පෙන්නුම් කරන්නේ පළමු කුරුල්ලන් තවමත් දුර්වල පියාසර කරන්නන් බවයි. පසුව පමණක්, පරිණාමයේ ක්රියාවලියේදී, සමස්ත සංවිධානයේ ක්රමයෙන් වැඩිදියුණු වීම හේතුවෙන්, කුරුල්ලන් පියාසර කිරීමට පරිපූර්ණ ලෙස අනුගත විය.
ක්ෂීරපායීන්ගේ සම්භවය
ක්ෂීරපායින් යනු පෘෂ්ඨවංශීන්ගේ ඉහළම පන්තිය වන අතර, උරගයින්, එනම් සත්ව-දත් කටුස්සන් සමඟ සම්භවය ද සම්බන්ධ වේ.
ක්ෂීරපායීන්ගේ පැරණිතම පොසිල නටබුන් මධ්ය යුගයේ ආරම්භයේ සිටම දැන සිටියහ.
ක්ෂීරපායින් ජීවිතයේ අරගලය ජය ගැනීමට අවස්ථාව ලබා දී ඇති බොහෝ ලක්ෂණ වලින් කැපී පෙනේ. ක්ෂීරපායීන්ගේ එවැනි ලක්ෂණ වන්නේ සීතලෙන්, මස්මින ග්රන්ථිවලින් මෙන්ම ගර්භාෂය තුළ පැටවුන් බිහිකිරීමෙන් ඔවුන් හොඳින් ආරක්ෂා කරන හිසකෙස් රේඛාවයි.
ක්ෂීරපායීන්ගේ උණුසුම් ලේ වැගිරීමේ ලක්ෂණය කුරුල්ලන් මෙන් ඔවුන්ට පැවැත්ම සඳහා වන අරගලයේදී විශාල වාසි ලබා දුන්නේය. ක්ෂීරපායීන්ගේ ප්රගතිශීලී පරිණාමය තුළ විශාල වැදගත්කමක් ද මොළයේ සැලකිය යුතු වර්ධනයකි.
මධ්යතන යුගය, සමහර විට "උරගයින්ගේ යුගය" ලෙස හඳුන්වනු ලබන අතර, කුරුල්ලන් සහ ක්ෂීරපායින් විසින් ආධිපත්යය දරන නව යුගයක් මගින් ප්රතිස්ථාපනය විය.
කේම්බ්රියන්හි ආත්රපෝඩාවන්ගේ පරිණාමය
භෞමික ආත්රපෝඩාවන් කණ්ඩායම් කිහිපයක නියෝජිතයන් ඩෙවෝනියන් වෙතින් හැඳින්වේ: පැලියෝසොයික් සන්නද්ධ මකුළුවන් (සොලූටා), මයිටාවන් (ඇකරොමෝර්ෆා) සහ පහළ ප්රාථමික පියාපත් රහිත කෘමීන් (කෘමිනාශක) කණ්ඩායම්. නිසැකව ම, ඩෙවෝනියානු යුගයේ භෞමික අපෘෂ්ඨවංශීන්ගේ විවිධත්වය අප වෙත පහළ වී ඇති කුඩා පොසිල අවශේෂවලින් යෝජනා කරන ලද ප්රමාණයට වඩා බෙහෙවින් වැඩි ය.
මුල් කාබොනිෆරස් යුගයේ දෙවන භාගයේදී, පියාපත් සහිත ඉහළ කෘමීන්, pterygotes, දර්ශනය විය. බොහෝ විට, පියාපත් සහිත කෘමීන්ගේ මුතුන් මිත්තන් ටන්ක සහ ගස් ඔටුනු මත කඳු නැගීමේ ජීවිතයකට මාරු විය. වැටීම් සහ පැනීමේදී ලිස්සා යාම සඳහා, මෙම සතුන් වෙහෙසකර කොටස් මත ශරීරයේ පාර්ශ්වීය පැතලි නෙරා යාම භාවිතා කළහ. මෙම නිශ්චල උපග්රන්ථ වලින්, අනුවර්තන පරිණාමයේ ක්රියාවලියේදී, ස්වාභාවික වරණය විශ්මයජනක ලෙස පරිපූර්ණ ගුවන් යානයක් නිර්මාණය කර ඇත. මුලදී, උරස් කොටස් 3 හි පියාපත් පැවතුනි, නමුත් ඉදිරිපස යුගලය තවමත් ස්ථාවර ගුවන් යානා ආකාරයෙන් ප්රාථමික ව්යුහයක් රඳවා තබා ඇත, උදාහරණයක් ලෙස, Paleodictyoptera (Palaeodictyoptera) (රූපය 48). දන්නා කෘමීන්ගේ කණ්ඩායම් බොහොමයක් මෙම ඉදිරිපස පියාපත් යුගලය අහිමි වී ඇත. Carboniferous අවසානය වන විට, පියාඹන කෘමීන්ගේ විවිධ කණ්ඩායම් දැනටමත් පැවතුනි, ඔවුන්ගෙන් සමහරක් වර්තමානය දක්වා නොනැසී පවතී (mayflies, dragonflies, cockroaches, orthoptera, homoptera, Scorpions). පැලියෝසොයික් යුගයේ කෘමීන් පමණක් පියාසර කරන සතුන් විය. සමහර මකරුන් විශේෂ පසුව විශාල ප්රමාණයට ළඟා විය: ඉහළ කාබොනිෆරස් - පහළ පර්මියන් මෙගනයුරා හි සිරුරේ දිග සෙන්ටිමීටර 30 දක්වා ළඟා විය, පියාපත් සෙන්ටිමීටර 65 දක්වා විය.
භූමිෂ්ඨ පෘෂ්ඨවංශීන්ගේ (tetrapods) මුතුන් මිත්තන් lobe-finned මසුන් වූ අතර, සැහැල්ලු හා බලවත් ලෙස මාංශ පේශී යුගල කළ අත් පා - sarcopterygium, මෙම මසුන්ට ඝන පඳුරු අතර ජලාශ පතුලේ ගමන් කිරීමට පමණක් නොව, නොගැඹුරු ලෙස පිටතට යාමට ඉඩ සලසයි. ජලය, එක් ජලාශයක සිට තවත් කුඩා නාලිකා ඔස්සේ බඩගා යන්න. මෙම අත් පා වල අභ්යන්තර ඇටසැකිල්ල සෑදී ඇත්තේ එක් විශාල අස්ථියකින් වන අතර, අත් පා වල ඉඟටිය (උරහිස් හෝ ශ්රෝණිය) සමඟ ප්රකාශ කර ඇති අතර, පසුව සමාන්තර අස්ථි දෙකක දෙවන කොටස වන අතර කුඩා අස්ථි ගණනාවක් දුරස්ථව පිහිටා ඇත (රූපය 45, c) . මෙම යෝජනා ක්රමය තුළ, භූමිෂ්ඨ පෘෂ්ඨවංශීන්ගේ අත් පා වල ඇටසැකිල්ලේ මූලාකෘතිය පහසුවෙන් දැකගත හැකිය.
නොගැඹුරු ජලයේ, වෙරළබඩ ශාක අතර සහ ජල කඳ ඉවුරුවල ජීවත් වූ අපෘෂ්ඨවංශීන්ගේ බහුල සත්ත්ව විශේෂ - සමහර lobe-finned මත්ස්ය විශේෂ නව ආහාර පදනමක් සංවර්ධනය කිරීම කරා ගමන් කර ඇත. අනුවර්තන පරිණාමයේ මෙම දිශාව තව දුරටත් විශාල විලෝපිකයන් ඇතුළත් නොවූ එහි වේගයෙන් වර්ධනය වන ශාක හා සත්ත්ව විශේෂ සමඟ ඉඩම් සංවර්ධනයට තවදුරටත් හේතු විය. මෙම පාරිසරික නිකේතනය භූමිෂ්ඨ පෘෂ්ඨවංශීන් විසින් අල්ලා ගන්නා ලදී.
පෘථිවි ජීවයට අනුවර්තනයන් ස්වාධීනව සහ සමාන්තරව lobe-finned මසුන්ගේ පරිණාමයේ රේඛා කිහිපයකින් වර්ධනය වූ බව සැලකිල්ලට ගත යුතුය. මේ සම්බන්ධයෙන්, E. Jarvik lobe-finned fish (Osteolepiformes සහ Porolepiformes) විවිධ කණ්ඩායම් දෙකකින් භෞමික පෘෂ්ඨවංශීන්ගේ diphyletic සම්භවය පිළිබඳ උපකල්පනයක් ඉදිරිපත් කළේය. කෙසේ වෙතත්, විද්යාඥයින් ගණනාවක් (A. Romer, I. I. Shmalgauzen, K. Thomson, E. I. Vorobyeva) Yarvik ගේ තර්ක විවේචනය කළහ. වර්තමානයේ, බොහෝ පර්යේෂකයන් ඔස්ටියෝලෙපිෆෝම් ක්රොසොප්ටෙරන් වලින් ටෙට්රාපොඩ් වල මොනොෆයිලටික් සම්භවය වඩාත් සම්භාවිතාවක් ලෙස සලකයි, නමුත් පැරෆිලියා ඇතිවීමේ හැකියාවට අවසර ඇත, i.e. සමාන්තරව පරිණාමය වූ ඔස්ටියෝලිපිෆෝම් මාළු වල සමීප සම්බන්ධිත ෆයිලෙටික් රේඛා කිහිපයක් මගින් උභයජීවීන්ගේ සංවිධානයේ මට්ටම සාක්ෂාත් කර ගැනීම.
උභයජීවීන්ගේ (පන්තියේ ඇම්ෆිබියා) ichthyostegi (Ichthyostegalia, Fig. 49) වඩාත්ම පැරණි නියෝජිතයන් ග්රීන්ලන්තයේ ඉහළ ඩෙවෝනියානු තැන්පතු වලින් හැඳින්වේ. Ichthyostegs තරමක් විශාල සතුන් (සෙන්ටිමීටර 80 ට වඩා දිග), ඒ වන විටත් හොඳින් වර්ධනය වී ඇති භූමිෂ්ඨ ආකාරයේ ඇඟිලි පහේ අත් පා තිබූ අතර, ඔවුන්ට නිසැකවම ගොඩබිමට බඩගා යා හැකිය. කෙසේ වෙතත්, Ichthyostegs බොහෝ විට ඔවුන්ගේ ජීවිතයේ සැලකිය යුතු කොටසක් ජල කඳවල ගත කළ අතර, Jarvik හි උචිත ප්රකාශනයට අනුව, "අඩි හතරේ මාළු" විය. ලොබ් වරල් සහිත මාළු වලිගයට සමාන සැබෑ පෞච්ඡ වරලක් තිබීම මෙන්ම වැඩිහිටි තත්වයේ භූ කම්පන සංවේදක ප්රතිග්රාහක සංරක්ෂණය කිරීම ("පාර්ශ්වික රේඛාව" - ජලජ පරිසරයේ පමණක් ක්රියා කරන සංවේදක ඉන්ද්රියයන් මෙයට සාක්ෂි දරයි. , අධෝරක්ත කම්පන සහ විද්යුත් චුම්භක ක්ෂේත්ර වටහා ගැනීම). Ichthyosteg හි හිස් කබලේ, ගිල් ආවරණයේ අස්ථිවල මූලයන් සංරක්ෂණය කර ඇත (සමහර විට, කාටිලේජිනස් ගිල් ආරුක්කු මත සවි කර ඇති ගිල් ද තිබිය හැකිය). සිරුර කුඩා අස්ථි කොරපොතු වලින් වැසී තිබුණි.
බොහෝ විට, ichthyostegas වැනි පැරණිතම උභයජීවීන්, ජලජ පරිසරයේ බොහෝ හා විවිධ lobe-finned මසුන් සමඟ තරඟය අහිමි විය, මන්ද යත්, භූමිෂ්ඨ ආකාරයේ යුගලනය කරන ලද අත් පා පිහිනීමේදී ඵලදායි ගැඹුර සුක්කානම් විය නොහැකි බැවිනි. ඇඟිලි පහේ අත් පා වල වාසි ප්රකාශ වූයේ ජල කඳේ සහ ගොඩබිමේ කුඩාම කොටස්වල පමණි. බොහෝ දුරට ඉඩ ඇත්තේ, ඩෙවෝනියන් හි ලූප් වරල් සහිත මසුන් සහ ඔවුන්ගෙන් පැවත එන උභයජීවීන් අතර තරඟකාරී සම්බන්ධතාවයක් ඇති අතර, එය මෙම කණ්ඩායම් විවිධ අනුවර්තන කලාපවලට අපසරනය වීමට දායක විය: ලූප් වරල් සහිත මසුන් ජල කඳවල ආධිපත්යය දරන අතර උභයජීවීන් අතරමැදි වාසය කළහ. ජලය සහ ගොඩබිම අතර වාසස්ථාන (කුඩා පිහිනීම දුෂ්කර; තෙත්බිම්; ගොඩබිමෙහි අධික තෙතමනය සහිත ප්රදේශ). පසුකාලීනව, මෙම වර්ගයේ වාසස්ථාන, ජලය සහ ගොඩබිම අතර අතරමැදි සහ මායිම්, අද දක්වා උභයජීවීන් සඳහා වඩාත්ම හිතකර අනුවර්ණ කලාපය ලෙස පවතින අතර, උභයජීවීන් මාළු සහ ටෙට්රාපොඩ් වල වඩාත් දියුණු භෞමික කණ්ඩායම් දෙකටම වඩා වාසි රඳවා ගෙන ඇති බව අපි සටහන් කරමු. අවසාන ඩෙවෝනියන් හි, පැරණිතම උභයජීවීන්ගේ අපසාරී පරිණාමය දැනටමත් ආරම්භ වී ඇත. Ichthyostegs වලට අමතරව, Devonian අවසානයේ තවත් උභයජීවී කණ්ඩායම් ගණනාවක නියෝජිතයන් සිටි අතර, ඒවායේ ෆොසිල අවශේෂ විවිධ ප්රදේශවලින් හමු විය. මෙම සතුන්ගේ ඇඟිලි ගණන බොහෝ විට 7-8 දක්වා ළඟා වීම කුතුහලයට කරුණකි.
Carboniferous දී, පුරාණ උභයජීවීන්ගේ මල් පිපීම ආරම්භ වන අතර, ඉහළ පැලියෝසොයික්හි විවිධාකාර ස්වරූප වලින් නියෝජනය වේ (රූපය 50), ඒවා ස්ටෙගෝසෙෆල්ස් නමින් කොන්දේසි සහිතව ඒකාබද්ධ වේ, i.e. ආවරණ හිස්. දැන් වර්ගීකරණ අර්ථයක් නොමැති මෙම යෙදුම, නාස්පුඩු, අක්ෂි කුහරවල විවරයන් මගින් පමණක් සිදුරු කරන ලද රයි (ඊනියා quilted හිස් කබල) වලින් උරුම වූ ඝන අස්ථි කවචයකින් හිස ආරක්ෂා කර ඇති මෙම සතුන්ගේ ලාක්ෂණික ලක්ෂණය අවධාරණය කරයි. සහ "parietal eye".
stegocephalians කණ්ඩායම් අතරින් වඩාත් ප්රසිද්ධ වන්නේ ichthyostegi ඇතුළත් labyrinthodonts (Labyrinthodontia) ය. ඔවුන්ගේ නම දත් වල ව්යුහයේ කුතුහලය දනවන ලක්ෂණයක් සමඟ සම්බන්ධ වී ඇති අතර එය වරල් සහිත මසුන්ගෙන් ද උරුම වේ: එනමල් සහ ඩෙන්ටින් බොහෝ නැමීම් සාදයි, එවිට තීර්යක් කොටසක ලිබ්රින්ටයකට සමාන රටාවක් දිස් වේ.
ඉහළ පැලියෝසොයික් හි Labyrinthodonts යනු පෘෂ්ඨවංශික කණ්ඩායම්වල වඩාත් පුලුල්ව පැතිරුනු සහ බහුල විශේෂයකි. මේවාට කුඩා හා විශාල, මීටර් 1.5 ට වැඩි ආකෘති ඇතුළත් විය. කාබොනිෆරස් වල ආධිපත්යය දැරුවේ දුර්වල අත් පා සහ දිගු ශරීරයක් ඇති විශේෂයන් වන අතර ඒවා බොහෝ වගුරු බිම්වල ජීවත් විය. Permian හි, විශාල පැතලි හිසක් සහිත විශාල කිඹුල් හැඩැති stegocephalians දර්ශනය විය (Eryops හා සමානව, Fig. 50, b බලන්න), මෙන්ම වඩා හොඳින් වර්ධනය වූ අත් පා සහ කෙටි වූ ශරීරය සහ වලිගය සහිත කුඩා විශේෂ (Cacops, Fig. 50, c) මෙම දෙවැන්න බොහෝ විට, ප්රධාන වශයෙන් ගොඩබිම, විවිධ අපෘෂ්ඨවංශීන් පෝෂණය කරමින් ජීවත් වූ නමුත්, සියලුම උභයජීවීන් මෙන්, ඔවුන්ට ඉහළ වායු ආර්ද්රතාවය සහ බිත්තර දැමීම සඳහා ජල කඳන් සමීපව අවශ්ය විය. විද්යාඥයන් ගණනාවකට අනුව, ජල කඳ ඉවුරු දිගේ තබාගෙන ජලයට පැන ගොඩබිම පදනම් වූ සතුරන්ගෙන් බේරුණු කුඩා ලබිරින්තොඩොන්ට්, අනුරන්ගේ (අනුර) මුතුන් මිත්තන් බවට පත් වූ අතර එය පර්මියන් යුගයේ අග භාගයේදී ඇති විය. මැඩගස්කරයේ මුල් ට්රයැසික් ප්රදේශයේ සිට වලිග රහිත උභයජීවී ට්රයඩෝබා-ට්රැචස්ගේ පැරණිතම නියෝජිතයා දැනටමත් කුඩා වලිගයක් රඳවාගෙන සිටියද ගෙම්බන්ගේ ලාක්ෂණික ශරීර සමානුපාතිකයන් සහ මූලික ව්යුහාත්මක ලක්ෂණ තිබුණි.
ෆොසිල තත්වයේ ඇති නවීන උභයජීවීන්ගේ තවත් අනුපිළිවෙලවල් දෙකක් පසුකාලීන තැන්පතු වලින් දනියි: මැද ජුරාසික් වලින් කෞඩේට් (උරෝඩෙලා), පාද රහිත හෝ සීසිලියන් (අපෝඩා) මුල් ජුරාසික් වලින්. ඔවුන්ගේ සම්භවය පිළිබඳ ගැටළුව තවමත් විවාදාත්මක ය. හිසෙහි ඇටසැකිල්ල සහ මාංශ පේශිවල බොහෝ සලකුනු අනුව, වලිග සහිත උභයජීවීන් අනුරන්ට සමීප වන අතර, සමහරවිට, ඔවුන් සමඟ පොදු සම්භවයක් ඇත.
නූතන උභයජීවීන් අතර බොහෝ පැතිවලින් වඩාත් ප්රාථමික ව්යුහය රඳවා ගනිමින් කකුල් රහිත උභයජීවීන් තරමක් දුරස්ව සිටියි. සමහර පාෂාණ විද්යාඥයින් ඔවුන්ගේ මූලාරම්භය ලෙපොස්පොන්ඩිලි (ලෙපොස්පොන්ඩිලි) උප පන්තියේ එක්සත් වූ ස්ටෙගෝසෙෆාලියන්ගේ ෆයිලොජෙනටික් කඳ සමඟ සම්බන්ධ කරයි. (තවත් දෘෂ්ටි කෝණයකට අනුව, නූතන උභයජීවීන්ගේ අනුපිළිවෙල තුනටම පොදු සම්භවයක් තිබුණි, බොහෝ විට labyrinthodonts වලින්).
ලෙපොස්පොන්ඩිලික් අතර, දැඩි ලෙස දිගටි ශරීරයක් සහ දුර්වල ලෙස වර්ධනය වූ අත් පා සහිත බොහෝ ආකාර තිබුණි. උදාහරණයක් ලෙස, ලයිසොරොෆ් (Lysorophia) වේ. ලෙපොස්පොන්ඩිල් හි තවත් අනුපිළිවෙලක නියෝජිතයින්ට - සිසිලියන් වැනි ස්ටෝර්ක්ස් (අයිස්ටෝපෝඩා) ඔවුන්ගේ අත් පා සම්පූර්ණයෙන්ම අහිමි විය. කෙසේ වෙතත්, ඔවුන්ට විශේෂිත ව්යුහාත්මක ලක්ෂණ ගණනාවක් තිබූ බැවින්, ඔවුන් සීසිලියානුවන්ගේ මුතුන් මිත්තන් විය නොහැක. Nectridia (Nectridia) ද lepospondylic අයත් වේ. ඔවුන්ගේ එක් පවුලක් හිස් කබලේ විකාර ව්යුහයකින් කැපී පෙනුණි, එහි පසුපස කොන් දිගු කේතුකාකාර වර්ධනයක් ලෙස අතිශයින් පසෙකට හා පසුපසට වැඩී ඇති අතර එමඟින් හිස සමස්තයක් ලෙස ඉහළින් බූමරංගයක් මෙන් දිස් විය. මෙම වර්ධනයට යම් ආකාරයක හයිඩ්රොෆොයිල් වල භූමිකාවක් ඉටු කළ හැකි යැයි උපකල්පනය කර ඇති අතර එමඟින් චලනය අතරතුර යම් සෝපානයක් නිර්මාණය වී ජලයේ ලිස්සා යාමට දායක වේ.
දැනටමත් ඩෙවෝනියන් හි, ප්රාථමික labyrinthodonts වලින් වෙන් කරන ලද ඇන්ත්රකෝසෝරයන් (ඇන්ත්රකොසෝරියා) කණ්ඩායමක්, එහි පැරණිතම නියෝජිතයා වන ටියුලර්පෙටන්, ටූලා කලාපයේ ඉහළ ඩෙවෝනියානු තැන්පතු වලින් සොයා ගන්නා ලදී. මුලදී, මෙම stegocephalians ද ජල කඳ වෙරළ තීරයේ ජීවත් වූහ. කෙසේ වෙතත්, ඇන්ත්රකෝසෝරයන් අතර, භෞමික වාසස්ථාන සංවර්ධනය කිරීමේ ප්රවණතාවක් ඉතා ඉක්මනින් දක්වා ඇත. බොහෝ අවස්ථාවන්හිදී මෙන්, මෙම ප්රවණතාවය ස්වාධීනව සහ සමාන්තරව විවිධ ෆයිලෙටික් පෙළපත් කිහිපයක වර්ධනය වූ අතර, ඒ සෑම එකක්ම ගොඩබිම ජීවිතයට බොහෝ දුරට සමාන අනුගතවීම් ක්රමයෙන් වර්ධනය විය. ඇන්ත්රකෝසෝරයන් සහ ඔවුන්ගෙන් පැවත එන්නන්, ඔවුන්ගේ භූමිෂ්ඨ ජීවන රටාව නොතකා, සාමාන්ය සංවිධානයේ මට්ටම සහ උභයජීවීන්ගේ ප්රජනනය සහ ඔන්ටොජෙනිස් වල ලාක්ෂණික ලක්ෂණ (ජල සිරුරු වල පැටවුන් සහ ජලජ කීටයන් සමඟ) උප පංතියකට ඒකාබද්ධ වේ. batrachosaurs, හෝ "Lizard-amphibians" ( Batrachosauria). පර්මියන් හි පැවති Seymouria (Seymouria; Fig. 51), Kot-lassia (Kotlassia) සහ ඒ ආශ්රිත ආකෘති batrachosaurs ට අයත් වේ. දිගු කලක් තිස්සේ, සේමූරියන් උභයජීවීන් නොව උරගයින් ලෙස සැලකේ, ඔවුන් හිස් කබලේ, කොඳු ඇට පෙළේ සහ අත් පා වල ව්යුහය තුළ සැබෑ උරගයින්ට එතරම් සමීප ය. කෙසේ වෙතත්, පසුව, මෙම සතුන්ගේ කීට අවධීන්ගේ නටබුන් සොයා ගන්නා ලද අතර, හිස් කබලේ අස්ථි මත භූ කම්පන සංවේදී අවයවවල නාලිකා වල අංශු මාත්ර තිබූ අතර, එබැවින් ජලයේ ජීවත් විය.
මුල් කාබොනිෆරස් හි සමහර ප්රාථමික බැට්රාචෝසෝරයන්ගෙන්, සැබෑ උරගයන් (උරගයින්) ඇති වූ අතර, විශේෂයෙන්, ස්කොට්ලන්තයේ පහළ කාබන් තැන්පතුවල දක්නට ලැබෙන වෙස්ට්ලෝතියානා හි කුඩා ස්වරූපයක් ඇතුළත් වේ. එහි සංවිධානය හිස් කබලේ ප්රගතිශීලී (උරග) ලක්ෂණ සහ කොඳු ඇට පෙළේ සහ අත් පා වල ("මොසෙයික්") ලක්ෂණ ගණනාවක ප්රාථමික තත්වය ඒකාබද්ධ කරයි. R. කැරොල් විශ්වාස කරන්නේ උරගයින්ගේ ආසන්නතම මුතුන් මිත්තන් ඔවුන්ගේ ප්රධාන ලක්ෂණ වල Late Carboniferous Gefirostegidae (Gcphyrostegidae, Fig. 51) ට සමීප විය හැකි බවයි, කෙසේ වෙතත්, උරගයින්ගේ සැබෑ මුතුන් මිත්තන් වීමට ප්රමාද වැඩියි. Gefirostegi යනු උරගයින්ගේ මුල් කාබොනිෆරස් මුතුන් මිත්තන්ගේ ලක්ෂණයක් වන පුරාවිද්යාත්මක ව්යුහාත්මක ලක්ෂණ ගණනාවක් රඳවා ගත් ධාතු සමූහයක් විය හැකිය.