مراحل اولیه جنین زایی. مراحل رشد جنین. سلول های جنسی مرد - اسپرم
جنین زایی (جنین یونانی - جنین، پیدایش - رشد) - دوره اولیه رشد فردی بدن از لحظه لقاح (لقاح) تا تولد، مرحله اولیه انتوژنز است (یونانی ontos - بودن، پیدایش - تکامل)، فرآیند رشد فردی بدن از لقاح تا مرگ.
رشد هر موجود زنده در نتیجه ادغام دو سلول جنسی (گامت) نر و ماده آغاز می شود. همه سلول های بدن، علی رغم تفاوت در ساختار و عملکرد، با یک چیز متحد می شوند - یک اطلاعات ژنتیکی ذخیره شده در هسته هر سلول، یک مجموعه واحد دوگانه از کروموزوم ها (به جز سلول های خونی بسیار تخصصی - گلبول های قرمز، که چنین نیستند. هسته دارند). یعنی تمام سلول های سوماتیک (سوما - بدن) دیپلوئید هستند و حاوی یک مجموعه کروموزوم دوگانه - 2 n هستند و فقط سلول های زایا (گامت ها) که در غدد جنسی تخصصی (بیضه ها و تخمدان ها) تشکیل می شوند حاوی یک مجموعه کروموزوم واحد هستند - 1. n
هنگامی که سلول های زاینده ادغام می شوند، یک سلول تشکیل می شود - یک زیگوت، که در آن یک مجموعه دوگانه از کروموزوم ها بازسازی می شود. به یاد بیاورید که هسته یک سلول انسانی به ترتیب دارای 46 کروموزوم است، سلول های زایا دارای 23 کروموزوم هستند.
زیگوت حاصل شروع به تقسیم می کند. اولین مرحله تقسیم زیگوت را خرد کردن می نامند که در نتیجه آن ساختار چند سلولی مورولا (توت) تشکیل می شود. سیتوپلاسم به طور نابرابر بین سلول ها توزیع می شود، سلول های نیمه پایینی مورولا بزرگتر از سلول های بالایی هستند. از نظر حجم، مورولا با حجم زیگوت قابل مقایسه است.
در مرحله دوم تقسیم، در نتیجه توزیع مجدد سلول ها، یک جنین تک لایه تشکیل می شود - بلاستولا، متشکل از یک لایه سلول و یک حفره (بلاستوکول). اندازه سلول های بلاستولا متفاوت است.
در مرحله III، به نظر می رسد سلول های قطب پایین به سمت داخل برآمدگی (تخلخل) دارند و یک جنین دو لایه تشکیل می شود - گاسترولا، متشکل از لایه بیرونی سلول ها - اکتودرم و لایه داخلی سلول ها - اندودرم.
خیلی زود در بین لایه های I و II سلول ها در اثر تقسیم سلولی لایه دیگری از سلول ها تشکیل می شود که وسط آن مزودرم است و جنین سه لایه می شود. این مرحله گاسترولا را کامل می کند.
از این سه لایه سلولی (که به آنها لایه های جوانه می گویند) بافت ها و اندام های موجود زنده آینده تشکیل می شود. از اکتودرم، بافت پوششی و عصبی رشد می کند، از مزودرم - اسکلت، ماهیچه ها، سیستم گردش خون، اندام های تناسلی، اندام های دفعی، از اندودرم - تنفس، تغذیه، کبد، پانکراس. بسیاری از اندام ها از چندین لایه جوانه تشکیل می شوند.
جنین زایی شامل فرآیندهای لقاح تا تولد است.
رشد بدن انسان پس از لقاح سلول زایای زن - تخمک (تخمک) مرد - اسپرم (اسپرم، اسپرم) آغاز می شود.
مطالعه دقیق رشد جنین انسان (جنین) موضوع جنین شناسی است. در اینجا ما فقط به یک مرور کلی از رشد جنین (جنین زایی) که برای درک بدن انسان ضروری است محدود می کنیم.
جنین زایی همه مهره داران از جمله انسان را می توان به سه دوره تقسیم کرد.
1. شکاف: یک تخمک بارور شده، اسپرمویوم یا زیگوت به طور متوالی به سلول ها (2،4،8،16 و غیره) تقسیم می شود، در نتیجه ابتدا یک توپ چند سلولی متراکم، مورولا، و سپس یک تک تشکیل می شود. وزیکول لایه - بلاستولا که حاوی حفره اولیه بلاستوکول است. مدت این دوره 7 روز می باشد.
2. گاسترولاسیون شامل تبدیل جنین تک لایه به دو لایه و بعداً سه لایه - گاسترولا است. دو لایه اول سلول ها را لایه های جوانه می نامند: اکتودرم بیرونی و اندودرم داخلی (تا دو هفته پس از لقاح) و سومین لایه میانی که بعداً بین آنها ظاهر می شود لایه میانی جوانه - مزودرم نامیده می شود. دومین نتیجه مهم گاسترولاسیون در همه آکوردها، پیدایش مجموعه محوری ابتدایی است: در سمت پشتی (پشتی) اندودرم، پایه رشته پشتی، وتر، و در سمت شکمی (شکمی) آن ظاهر می شود. پایه اندودرم روده؛ در سمت پشتی جنین، در امتداد خط وسط آن، صفحه عصبی از اکتودرم - پایه سیستم عصبی آزاد می شود، و بقیه اکتودرم به ساخت اپیدرم پوست می رود و به همین دلیل اکتودرم پوست نامیده می شود.
متعاقباً جنین رشد می کند و به شکل استوانه ای با انتهای دمی سر (جمجمه ای) و دمی تبدیل می شود. این دوره تا پایان هفته سوم پس از لقاح ادامه دارد.
3. اندام زایی و هیستوژنز: صفحه عصبی در زیر اکتودرم فرو می رود و به یک لوله عصبی تبدیل می شود که از بخش های جداگانه - نوروتوم ها - تشکیل شده و باعث رشد سیستم عصبی می شود. پایه های مزودرمی از اندودرم روده اولیه جدا شده و یک ردیف از کیسه های متامریک تشکیل می دهند که در کناره های بدن جنین رشد می کنند و هر کدام به دو بخش تقسیم می شوند: پشتی که در طرفین قرار دارد. نوتوکورد و لوله عصبی و شکم که در دو طرف بدن جنین قرار دارد. بخشهای پشتی مزودرم بخشهای اولیه بدن - سومیتها را تشکیل میدهند که هر کدام به نوبه خود به یک اسکلروتوم تقسیم میشوند که باعث ایجاد اسکلت و یک میوتوم میشود که از آن ماهیچهها رشد میکنند. از سومیت (در سمت جانبی آن)، یک بخش پوست نیز متمایز می شود - یک درماتوم. بخشهای شکمی مزودرم که اسپلانکنوتوم نامیده میشوند، کیسههای جفتی را تشکیل میدهند که حاوی حفره ثانویه بدن هستند.
اندودرم روده که پس از جدا شدن نوتوکورد و مزودرم باقی مانده است، روده ثانویه را تشکیل می دهد - اساس رشد اندام های داخلی. متعاقباً تمام اندام های بدن گذاشته می شوند که مواد ساخت آنها سه لایه جوانه است.
1. از لایه بیرونی جوانه، اکتودرم، توسعه می یابد:
الف) اپیدرم پوست و مشتقات آن (مو، ناخن، غدد پوست).
ب) اپیتلیوم غشای مخاطی بینی، دهان و مقعد.
ج) سیستم عصبی و اپیتلیوم اندام های حسی.
2. از لایه داخلی زایا، اندودرم، اپیتلیوم مخاط بیشتر دستگاه گوارش با تمام ساختارهای غده ای متعلق به آن، بیشتر اندام های تنفسی و همچنین اپیتلیوم تیروئید و غدد گواتر ایجاد می شود. .
3. از لایه میانی جوانه، مزودرم، ماهیچه های اسکلت، مزوتلیوم غشای حفره های سروزی با ابتدایی غدد جنسی و کلیه ها ایجاد می شود.
علاوه بر این، بافت همبند جنینی، مزانشیم، از بخشهای پشتی مزودرم ایجاد میشود که باعث ایجاد انواع بافت همبند، از جمله غضروف و استخوان میشود. از آنجایی که در ابتدا مزانشیم مواد مغذی را به قسمت های مختلف جنین هدایت می کند و عملکرد تغذیه ای را انجام می دهد، بعداً خون، لنف، رگ های خونی، غدد لنفاوی و طحال از آن ایجاد می شود.
علاوه بر رشد خود جنین، باید به شکل گیری قسمت های خارج از جنین نیز توجه شود که با کمک آنها جنین مواد مغذی لازم برای زندگی خود را دریافت می کند.
در یک توپ متراکم چند سلولی، یک ندول ژرمینال داخلی، یک جنین و یک لایه بیرونی از سلول ها متمایز می شود که نقش مهمی در تغذیه جنین ایفا می کند و به همین دلیل تروفوبلاست نامیده می شود. با کمک تروفوبلاست، جنین به ضخامت غشای مخاطی رحم نفوذ می کند (لانه گزینی) و در اینجا تشکیل اندام خاصی آغاز می شود که به کمک آن جنین با بدن مادر و تغذیه آن مرتبط می شود. انجام می شود. این اندام جای نوزاد، بستر یا جفت نامیده می شود. به پستاندارانی که جفت دارند جفت می گویند. در کنار تشکیل جفت، روند جداسازی جنین وجود دارد که از قسمت های خارج جنینی در نتیجه ظهور چین به اصطلاح تنه ایجاد می شود که با یک برآمدگی به وسط می رود. به نظر می رسد که بدن جنین را از قسمت های خارج جنینی با یک حلقه می بندد. با این حال، در همان زمان، ارتباط با جفت با کمک ساقه ناف حفظ می شود، که سپس به بند ناف تبدیل می شود. در مراحل اولیه رشد، مجرای زرده از طریق دومی عبور می کند که روده را با بیرون زدگی آن به ناحیه خارج جنینی، کیسه زرده، متصل می کند. در مهره دارانی که جفت ندارند، کیسه زرده حاوی مواد مغذی تخم مرغ - زرده است و اندام مهمی است که جنین از طریق آن تغذیه می شود.
در انسان کیسه زرده اگرچه ایجاد می شود اما نقش چندانی در رشد جنین ندارد و پس از جذب محتویات آن به تدریج کاهش می یابد. عروق ناف (جفت) نیز از طریق بند ناف عبور می کنند، که از طریق آن خون از جفت به بدن جنین و عقب جریان می یابد. آنها از مزودرم کیسه ادراری یا آلانتویس که از دیواره شکمی روده بیرون زده و از بدن جنین از طریق دهانه ناف به قسمت خارج جنینی خارج می شود، ایجاد می شوند. در انسان، بخشی از آلانتویس که در وسط بدن جنین قرار دارد، بخشی از مثانه را تشکیل میدهد و رگهای خونی ناف از رگهای آن تشکیل میشوند. جنین در حال رشد با دو غشای زاینده پوشیده شده است. غشای داخلی، آمنیون، یک کیسه حجیم تشکیل می دهد که با مایع پروتئینی پر شده و یک محیط مایع برای جنین تشکیل می دهد که از طریق آن کیسه، غشای آبی نامیده می شود. کل جنین، همراه با کیسه های آمنیوتیک و زرده، توسط یک غشای خارجی (که شامل تروفوبلاست نیز می شود) احاطه شده است. این پوسته با داشتن پرز، پرز یا کوریون نامیده می شود. کوریون عملکردهای تغذیه ای، تنفسی، دفعی و مانع را انجام می دهد.
13. برخلاف خزهها، در سرخسها، دم اسبها و خزههای کلوبی، اسپوروفیت، گیاهی برگدار، در چرخه رشد غالب است. نمایندگان این سه گروه از گیاهان دارای برگ، ساقه و ریشه هستند. اکثر آنها دارای ریزوم های زیرزمینی با برگ های تغییر یافته و ریشه های ناخواسته هستند. دم اسبهای مدرن، خزههای کلوبی و سرخسها عمدتاً گیاهانی علفی هستند. فقط در مناطق گرمسیری و نیمه گرمسیری سرخس درختی وجود دارد. با این حال، در دوران باستان - 200-350 میلیون سال پیش، این گروه از گیاهان با اشکال درخت مانند نشان داده می شدند و جنگل های انبوهی را تشکیل می دادند که باعث ایجاد بزرگترین سنگ شد.
ذخایر زغال سنگ جهان (Donbass، Kuzbass و غیره).
ویژگی های ساختاری خزه های باشگاهی، دم اسبی و سرخس چیست؟
ویژگی های خزه های باشگاهی، دم اسبی و سرخس ها را در نظر بگیرید. لیکوپسیدهای مدرن چند ساله و معمولاً همیشه سبز هستند. معروف ترین نماینده خزه های باشگاهی خزه های معمولی است که در روسیه مرکزی در صنوبر مرطوب و رایج است
جنگل های کاج این گیاه با ساقه ای منعطف و منشعب است که در امتداد زمین می خزند. برگها کوچک هستند و به صورت مارپیچی روی ساقه قرار گرفته اند. در اواخر تابستان معمولاً دو سنبلچه هاگ دار در شاخه های کناری ظاهر می شود. هر سنبلچه توسط برگهای کوچک، نازک و اصلاح شده به نام اسپوروفیل تشکیل می شود. در قاعده اسپوروفیل ها اسپورانژیا وجود دارد که در آن هاگ ها تشکیل می شود. دم اسبی یا دم اسبی با ساختار تقسیم شده ساقه ها به راحتی قابل تشخیص است: آنها دارای تناوب مشخصی از گره ها و میانگره ها هستند. برگ های روی ساقه به صورت حلقه هایی (چند قطعه در هر گره) قرار گرفته اند که ساقه را احاطه کرده اند. در بالای ساقه ها، سنبلچه های اسپوردار تشکیل می شود که در آنها هاگ ها می رسند. در برخی از گونه ها، به عنوان مثال، در دم اسب، ساقه ها دو نوع است: هاگ دار (صورتی مایل به قهوه ای، در بهار رشد می کند و پس از هاگ زایی می میرد) و رویشی (در تابستان از همان ریزوم ظاهر می شود). سرخس ها در طبیعت با گیاهان چند ساله، انگور، درختان و اپی فیت هایی که روی تنه درختان می نشینند نشان داده می شوند. سرخس ها برگ های بزرگی دارند. بچه ها معمولاً مانند حلزون حلقه می شوند. سرخس های کشور ما دارای ریزوم هستند. اسپورانژیوم آنها در قسمت زیرین برگ قرار دارد و به صورت انبوه جمع آوری می شود - به آنها سوری می گویند. سرخس های منقرض شده دارای اسپورانژیای منفرد هستند. در قلمرو کشور ما سرخس هایی مانند براکن معمولی، علف اسب نر، بولتوس ماده، صدپا معمولی و گونه های دیگر وجود دارد.
ویژگی رشد سرخس ها، دم اسب ها و خزه های کلاب چیست؟
تولیدمثل در هر سه گروه از گیاهان اسپور بالاتر طبق یک طرح انجام می شود. آن را به عنوان مثال یک سرخس در نظر بگیرید. در سطح زیرین برگ گیاه بالغ، اسپورانژیوم با هاگ ایجاد می شود. هنگامی که در شرایط مساعد قرار گرفت، هاگ جوانه می زند و گامتوفیت را ایجاد می کند. به نظر می رسد یک صفحه کوچک با ریزوئید است و به آن رشد می گویند. رشد گامتانژی نر و ماده با
سلول های جنسی - تخمک و اسپرم. پس از لقاح، که در حضور آب اتفاق می افتد، جنین ابتدا از زیگوت، و سپس گیاه بالغ - اسپوروفیت رشد می کند. بنابراین، در خزه های باشگاهی، دم اسب ها و سرخس ها، یک تناوب از نسل جنسی (برآمدگی - گامتوفیت) و غیر جنسی (بالغ) وجود دارد.
گیاه لو - اسپوروفیت).
14. رشد پس از جنین: مستقیم و غیر مستقیم. دلایل تضعیف رقابت بین والدین و فرزندان در رشد غیر مستقیم.
1. رشد فردی ارگانیسم (آنتوژنز) - دوره زندگی که در طی تولید مثل جنسی با تشکیل زیگوت شروع می شود، با تغییرات برگشت ناپذیر (افزایش جرم، اندازه، ظاهر بافت ها و اندام های جدید) مشخص می شود. با مرگ به پایان می رسد
2. دوره های جنینی (جنینی) و پس از جنینی (پس از جنینی) رشد فردی ارگانیسم.
3. رشد پس از جنین (جایگزین جنین می شود) - دوره از تولد یا خروج جنین از تخم تا مرگ. راه های مختلف رشد پس از جنینی حیوانات - مستقیم و غیر مستقیم:
1) رشد مستقیم - تولد فرزندانی که شبیه یک ارگانیسم بالغ است. به عنوان مثال: رشد ماهی، خزندگان، پرندگان، پستانداران، برخی از انواع حشرات. بنابراین، یک ماهی کوچک شبیه یک ماهی بالغ، یک جوجه اردک شبیه یک اردک، یک بچه گربه شبیه یک گربه است.
2) رشد غیرمستقیم - تولد یا خروج از تخم فرزندان که از نظر خصوصیات مورفولوژیکی، سبک زندگی (نوع تغذیه، ماهیت حرکت) با ارگانیسم بالغ متفاوت است. به عنوان مثال: از تخم های خروس، لاروهای کرم مانند ظاهر می شوند، بر خلاف یک سوسک بالغ (روی درخت زندگی می کند، از برگ ها تغذیه می کند) در خاک زندگی می کنند و از ریشه تغذیه می کنند.
مراحل رشد غیر مستقیم حشرات: تخم، لارو، شفیره، بالغ. ویژگی های زندگی حیوانات در مرحله تخم و شفیره - آنها بی حرکت هستند. سبک زندگی فعال لارو و ارگانیسم بالغ، شرایط مختلف زندگی، استفاده از مواد غذایی مختلف.
4. اهمیت توسعه غیرمستقیم تضعیف رقابت بین والدین و فرزندان است، زیرا آنها غذاهای مختلف می خورند، زیستگاه های متفاوتی دارند. توسعه غیرمستقیم انطباق مهمی است که در فرآیند تکامل به وجود آمد. این به تضعیف مبارزه برای وجود بین والدین و فرزندان، بقای حیوانات در مراحل اولیه رشد پس از جنین کمک می کند.
خصوصیات عمومی اولین ژیمنوسپرم ها در پایان دوره دونین در حدود 350 میلیون سال پیش ظاهر شدند. آنها احتمالاً از سرخس های باستانی که در آغاز دوره کربونیفر منقرض شده اند، منشاء گرفته اند. در عصر مزوزوئیک - عصر کوهسازی، ظهور قارهها و خشک شدن آب و هوا - ژیمنوسپرمها به اوج خود رسیدند، اما قبلاً از اواسط کرتاسه موقعیت غالب خود را به آنژیوسپرمها از دست دادند.
بخش ژیمنوسپرم های مدرن شامل بیش از 700 گونه است. علیرغم تعداد نسبتاً کم گونه ها، ژیمنوسپرم ها تقریباً کل کره زمین را تسخیر کرده اند. در عرض های جغرافیایی معتدل نیمکره شمالی، جنگل های مخروطی به نام تایگا را در پهنه های وسیع تشکیل می دهند.
ژیمنوسپروم های مدرن عمدتاً توسط درختان، بسیار کمتر توسط درختچه ها و به ندرت توسط لیانا نشان داده می شوند. هیچ گیاه علفی در بین آنها وجود ندارد. برگهای ژیمنوسپرم نه تنها از نظر شکل و اندازه، بلکه از نظر مورفولوژی و آناتومی نیز با سایر گروههای گیاهان متفاوت است. در اکثر گونه ها، آنها سوزنی (سوزن) یا پوسته پوسته هستند. در برخی از نمایندگان، آنها بزرگ هستند (به عنوان مثال، در Velvichia شگفت انگیز، طول آنها به 2-3 متر می رسد)، به صورت پینه بریده، دولوبی، و غیره.
اکثریت قریب به اتفاق ژیمنوسپرم ها گیاهان همیشه سبز، تک یا دوپایه با ساقه و سیستم ریشه ای توسعه یافته هستند که توسط ریشه های اصلی و جانبی تشکیل شده است. آنها توسط دانه هایی که از تخمک ها تشکیل می شوند پخش می شوند. تخمک ها برهنه هستند (از این رو نام این بخش) روی مگاسپوروفیل ها یا روی فلس های دانه جمع آوری شده در مخروط های ماده قرار دارند.
در چرخه رشد ژیمنوسپرم ها، یک تغییر متوالی از دو نسل وجود دارد - اسپوروفیت و گامتوفیت با غلبه اسپوروفیت. گامتوفیت ها به شدت کاهش می یابند و گامتوفیت های نر هولو و آنژیوسپرم ها آنتریدیا ندارند که به شدت با همه گیاهان بی دانه هتروسپور متفاوت است.
Gymnosperms شامل شش طبقه است که دو دسته از آنها کاملاً ناپدید شده اند و بقیه توسط گیاهان زنده نشان داده شده اند. حفظشدهترین و پرتعدادترین گروه از اسپرمها کلاس مخروطیها هستند که حداقل 560 گونه را شامل میشود که جنگلهایی را در پهنههای وسیع اوراسیا شمالی و آمریکای شمالی تشکیل میدهند. بیشترین تعداد گونه های کاج، صنوبر، کاج اروپایی در امتداد سواحل اقیانوس آرام یافت می شود.
کلاس سوزنی برگ. تمام مخروطیان درختان یا درختچه هایی همیشه سبز و به ندرت برگریز (مثلاً کاج اروپایی) با برگ های سوزنی یا فلس مانند (مثلاً سرو) هستند. برگ های سوزنی شکل (سوزن) متراکم، چرمی و سخت هستند که با لایه ای ضخیم از کوتیکول پوشیده شده اند. روزنه ها در فرورفتگی های پر از موم غوطه ور می شوند. همه این ویژگی های ساختار برگ ها سازگاری خوبی برای درختان مخروطی برای رشد در هر دو زیستگاه خشک و سرد فراهم می کند.
درختان مخروطی دارای تنه های ایستاده ای هستند که با پوسته پوسته پوسته پوشیده شده اند. در قسمت عرضی ساقه، چوب توسعه یافته و پوست و مغز کمتر توسعه یافته به وضوح قابل مشاهده است. آوند چوبی مخروطیان 90-95 درصد توسط تراکئیدها تشکیل شده است. مخروط های مخروطی دوپایه هستند. گیاهان - اغلب یک پایه، کمتر - دوپایه.
گسترده ترین نمایندگان مخروطیان در بلاروس و روسیه کاج اسکاچ و صنوبر نروژ یا اروپایی هستند. ساختار، تولید مثل، تناوب نسل آنها در چرخه توسعه نشان دهنده ویژگی های مشخصه همه مخروطیان است.
کاج اسکاچ یک گیاه تک پایه است (شکل 9.3). در ماه مه، دسته هایی از مخروط های نر زرد مایل به سبز به طول 4-6 میلی متر و قطر 3-4 میلی متر در پایه شاخه های کاج جوان تشکیل می شوند. در محور چنین مخروطی، برگ های فلس دار چندلایه یا میکروسپوروفیل ها قرار دارند. در سطح پایینی میکروسپوروفیل ها دو میکروسپورانژی وجود دارد - کیسه های گرده که در آنها گرده تولید می شود. هر دانه گرده دارای دو کیسه هوا است که حمل گرده توسط باد را آسان تر می کند. دو سلول در دانه گرده وجود دارد که یکی از آنها متعاقباً با برخورد به تخمک یک لوله گرده تشکیل می دهد و دیگری پس از تقسیم دو اسپرم تشکیل می دهد.
روی شاخه های دیگر همان گیاه، مخروط های ماده مایل به قرمز تشکیل می شود. در محور اصلی آنها فلس های پوششی شفاف کوچکی وجود دارد که در زیر بغل آن ها فلس های بزرگ ضخیم و متعاقباً چسبیده قرار دارند. در سمت بالایی این فلس ها دو تخمک وجود دارد که هر یک از آنها یک گامتوفیت ماده ایجاد می کند - یک آندوسپرم با دو آرکگونیا با یک تخم بزرگ در هر یک از آنها. در بالای تخمک، که از خارج توسط پوشش محافظت می شود، یک سوراخ وجود دارد - ورودی گرده، یا میکروپیل.
در اواخر بهار یا اوایل تابستان، گرده های بالغ توسط باد حمل می شوند و روی تخمک می ریزند. از طریق میکروپیل، گرده به داخل تخمک کشیده میشود، جایی که به داخل لوله گرده رشد میکند که به آرکگونیا نفوذ میکند. دو اسپرم تشکیل شده در این زمان از طریق لوله گرده به آرکگونیوم می روند. سپس یکی از اسپرم ها با تخمک ادغام می شود و دیگری می میرد. از یک تخم بارور شده (زیگوت) یک جنین بذری تشکیل می شود و تخمک به دانه تبدیل می شود. دانههای کاج در سال دوم میرسند، از مخروطها بیرون میریزند و توسط حیوانات یا باد جمعآوری میشوند و در فواصل قابل توجهی حمل میشوند.
از نظر اهمیت در زیست کره و نقش آنها در فعالیت های اقتصادی انسان، مخروطیان پس از آنژیوسپرم ها جایگاه دوم را به خود اختصاص داده اند و به مراتب از سایر گروه های گیاهان عالی پیشی گرفته اند.
آنها به حل مشکلات عظیم حفاظت از آب و چشم انداز کمک می کنند، به عنوان مهم ترین منبع چوب، مواد خام برای تولید کلوفون، سقز، الکل، مومیایی کردن، روغن های ضروری برای صنعت عطر، دارویی و سایر مواد ارزشمند عمل می کنند. برخی از درختان مخروطی به عنوان زینتی کشت می شوند (صنوبر، درخت درخت، سرو، سرو و غیره). دانه های تعدادی کاج (سیبری، کره ای، ایتالیایی) خورده می شود، روغن نیز از آنها به دست می آید.
نمایندگان سایر طبقات ژیمنوسپرم ها (سیکاد، گنتس، جینکو) بسیار نادرتر و کمتر از مخروطیان شناخته شده هستند. با این حال، تقریباً همه انواع سیکاد تزئینی هستند و به طور گسترده ای در بین باغبانان در بسیاری از کشورها محبوب هستند. درختچه های کم برگ همیشه سبز افدرا (کلاس گنتا) به عنوان منبع مواد خام برای تولید آلکالوئید افدرین است که به عنوان وسیله ای برای تحریک سیستم عصبی مرکزی و همچنین در درمان بیماری های آلرژیک استفاده می شود.
16. تقارن شعاعی- شکلی از تقارن که در آن یک جسم (یا شکل) با خود منطبق است وقتی یک جسم به دور یک نقطه یا خط خاص می چرخد. اغلب این نقطه با مرکز تقارن جسم، یعنی نقطه ای که در آن تعداد نامحدودی از محورهای تقارن دو طرفه قطع می شود، منطبق است. تقارن شعاعی توسط اجسام هندسی مانند دایره، توپ، استوانه یا مخروط وجود دارد.
در زیست شناسی، زمانی که یک یا چند محور تقارن از یک موجود سه بعدی عبور می کند، از تقارن شعاعی صحبت می شود. علاوه بر این، حیوانات متقارن شعاعی ممکن است صفحات تقارن نداشته باشند. بنابراین، سیفونوفور Velella دارای یک محور تقارن مرتبه دوم و بدون صفحات تقارن است.
معمولاً دو یا چند صفحه تقارن از محور تقارن عبور می کنند. این صفحات در یک خط مستقیم - محور تقارن - متقاطع می شوند. اگر حیوان به میزان معینی حول این محور بچرخد، روی خودش نمایش داده می شود (مطابق با خودش).
می تواند چندین محور تقارن (تقارن پلی اکسون) یا یکی (تقارن مونواکسون) وجود داشته باشد. تقارن پلیاکسون در میان پروتیست ها (مانند رادیولاریان ها) رایج است.
به عنوان یک قاعده، در حیوانات چند سلولی، دو انتهای (قطب) یک محور تقارن یکسان نیستند (به عنوان مثال، در چتر دریایی، دهان روی یک قطب (دهانی) و بالای زنگ در مقابل قرار دارد. (ابورال) چنین تقارنی (نوعی از تقارن شعاعی) در آناتومی مقایسه ای نامیده می شود در یک برجستگی دوبعدی، تقارن شعاعی را می توان در صورتی حفظ کرد که محور تقارن عمود بر صفحه برآمده باشد. به عبارت دیگر، حفظ تقارن شعاعی. بستگی به زاویه دید دارد
تقارن شعاعی مشخصه بسیاری از درختان خاردار و همچنین بیشتر خارپوستان است. در میان آنها به اصطلاح پنج تقارن وجود دارد که بر اساس پنج صفحه تقارن است. در خارپوستان، تقارن شعاعی ثانویه است: لاروهای آنها به صورت دو طرفه متقارن هستند، در حالی که در حیوانات بالغ، تقارن شعاعی خارجی با وجود صفحه مادرپوره نقض می شود.
علاوه بر تقارن شعاعی معمولی، تقارن شعاعی دو پرتو نیز وجود دارد (دو صفحه تقارن، به عنوان مثال، در ctenophores). اگر فقط یک صفحه تقارن وجود داشته باشد، تقارن دو طرفه است (حیوانات گروه بیلاتریا چنین تقارنی دارند).
در گیاهان گلدار، گلهای متقارن شعاعی اغلب یافت می شود: 3 صفحه تقارن (شاهی قورباغه ای)، 4 صفحه تقارن (Potentilla مستقیم)، 5 صفحه تقارن (گل زنگوله)، 6 صفحه تقارن (colchicum). گلهای با تقارن شعاعی را اکتیومورف و گلهای دارای تقارن دو طرفه را زیگومورف می نامند.
تقارن دو طرفه(تقارن دو طرفه) - تقارن انعکاس آینه که در آن جسم دارای یک صفحه تقارن است که دو نیمه آن متقارن آینه هستند. اگر یک عمود بر روی صفحه تقارن از نقطه A پایین بیاید و سپس از نقطه O در صفحه تقارن به طول AO کشیده شود، آنگاه به نقطه A1 می افتد که در همه چیز شبیه به نقطه A است. بدون محور تقارن برای اجسام متقارن دو طرفه. در حیوانات، تقارن دو طرفه در شباهت یا هویت تقریباً کامل نیمه چپ و راست بدن آشکار می شود. در این مورد، همیشه انحرافات تصادفی از تقارن وجود دارد (به عنوان مثال، تفاوت در خطوط پاپیلری، انشعاب عروق و محل خال در دست راست و چپ یک فرد). اغلب تفاوتهای کوچک اما منظم در ساختار خارجی (مثلاً ماهیچههای توسعهیافتهتر دست راست در افراد راستدست) و تفاوتهای مهمتر بین سمت راست و چپ بدن در محل اندامهای داخلی وجود دارد. به عنوان مثال، قلب در پستانداران معمولاً به صورت نامتقارن قرار می گیرد، با تغییر به چپ.
در حیوانات، ظهور تقارن دو طرفه در تکامل با خزیدن در امتداد بستر (در امتداد کف مخزن) همراه است که در ارتباط با آن قسمت پشتی و شکمی و همچنین نیمه راست و چپ بدن ظاهر می شود. به طور کلی، در میان حیوانات، تقارن دو طرفه در اشکال فعالانه متحرک بیشتر از شکل های بی حرکت است. تقارن دو طرفه مشخصه همه حیوانات به اندازه کافی سازمان یافته است، به جز خارپوستان. در سایر پادشاهی های موجودات زنده، تقارن دو طرفه مشخصه تعداد کمتری از اشکال است. در میان پروتیست ها، مشخصه دیپلمونادها (به عنوان مثال، ژیاردیا)، برخی از اشکال تریپانوزوم ها، بودونیدها و پوسته های بسیاری از روزن داران است. در گیاهان، تقارن دو طرفه معمولاً کل ارگانیسم نیست، بلکه بخش های جداگانه آن - برگ یا گل است. از نظر گیاه شناسی، گل های متقارن دو طرفه را زیگومورف می نامند.
17. آنژیوسپرم (گلدار، مادگی)با توجه به زمان ظهور آنها بر روی زمین، آنها جوان ترین و در عین حال سازمان یافته ترین گروه گیاهان هستند. در روند تکامل، نمایندگان این بخش دیرتر از دیگران ظاهر شدند، اما آنها خیلی سریع موقعیت غالب در جهان را اشغال کردند.
بارزترین ویژگی متمایز آنژیوسپرم ها وجود یک اندام عجیب و غریب در آنها است - گلی که در نمایندگان سایر بخش های گیاهی وجود ندارد. بنابراین، گیاه گلدان اغلب گیاهان گلدار نامیده می شود. تخمک آنها پنهان است، در داخل مادگی، در تخمدان آن رشد می کند، بنابراین آنژیوسپروم ها در غیر این صورت مادگی نامیده می شوند. گرده گلهای آنژیوسپرمها نه با تخمکها، مانند ژیمنوسپرمها، بلکه توسط یک تشکیل خاص - یک کلاله که به یک مادگی ختم میشود، گرفته میشود.
پس از لقاح تخمک، تخمک به دانه تبدیل می شود و تخمدان به میوه تبدیل می شود. در نتیجه، دانههای موجود در گل نباتی در میوهها رشد میکنند، به همین دلیل است که این بخش از گیاهان آنژیوسپرم نامیده میشود.
Angiospermae (Angiospermae) یا گل دهنده (Magnoliophyta) - بخش کامل ترین گیاهان عالی که دارای گل هستند. قبلاً آنها به همراه ژیمنوسپرم ها در بخش گیاهان بذر قرار می گرفتند. بر خلاف دومی، تخمک های گلدار در تخمدانی محصور شده اند که از برچه های جوش خورده تشکیل شده است.
گل اندام مولد آنژیوسپرم ها است. از یک پایه و یک ظرف تشکیل شده است. روی دومی پریانت (ساده یا دوتایی)، آندروسیوم (مجموعه ای از برچه ها) و ژینوسیوم (مجموعه ای از برچه ها) قرار دارند. هر پرچم شامل یک رشته نازک و یک بساک منبسط شده است که در آن اسپرم بالغ می شود. برچه گیاهان گلدار با یک مادگی نشان داده می شود که از یک تخمدان بزرگ و یک سبک بلند تشکیل شده است که قسمت امتداد یافته رأسی آن کلاله نامیده می شود.
آنژیوسپرم ها دارای اندام های رویشی هستند که پشتیبانی مکانیکی، حمل و نقل، فتوسنتز، تبادل گاز و ذخیره مواد مغذی و اندام های مولد درگیر در تولید مثل جنسی را فراهم می کنند. ساختار داخلی بافت ها پیچیده ترین در بین همه گیاهان است. عناصر غربال آبکش توسط سلول های همراه احاطه شده است. تقریباً همه آنژیوسپروم ها دارای آوند چوبی هستند.
گامت های نر موجود در داخل دانه های گرده روی کلاله می افتند و جوانه می زنند. گامتوفیت های گلدار بسیار ساده و مینیاتوری هستند که به طور قابل توجهی مدت چرخه تولید مثل را کاهش می دهد. آنها در نتیجه حداقل تعداد میتوز (سه مورد در گامتوفیت ماده و دو مورد در نر) تشکیل می شوند. یکی از ویژگیهای تولید مثل جنسی، لقاح مضاعف است، زمانی که یکی از اسپرمها با تخمک ترکیب میشود و یک زیگوت تشکیل میدهد و دومی با هستههای قطبی، آندوسپرم را تشکیل میدهد که به عنوان منبع مواد مغذی عمل میکند. دانه های گیاهان گلدار در یک میوه محصور شده اند (از این رو نام دوم آنها - گلداندان است).
18. DNAاین پدیده در آزمایش هایی با پنوموکوک ها، یعنی با باکتری هایی که باعث ذات الریه می شوند، کشف شد. دو شکل پنوموکوک شناخته شده است: فرم A با کپسول پلی ساکارید و فرم B بدون کپسول. هر دوی این صفات ارثی هستند.
پنوموکوک های A-form هنگامی که موش ها را آلوده می کنند باعث ذات الریه می شوند که در نتیجه موش ها می میرند. فرم B برای آنها بی ضرر است.
در سال 1928، باکتری شناس انگلیسی F. Griffiths موش ها را با مخلوطی از پنوموکوک های A-form که در اثر گرما کشته شده بودند و پنوموکوک B-form زنده را آلوده کرد. دانشمند فرض کرد که موش ها بیمار نمی شوند. اما بر خلاف انتظار، حیوانات آزمایشی مردند. F. Griffiths موفق شد پنوموکوک را از بافت موش های مرده جدا کند. معلوم شد که همه آنها کپسوله شده اند، یعنی فرم های A. در نتیجه، شکل کشته شده به نوعی خواص خود را به سلول های زنده فرم B منتقل کرد. اما چگونه؟ با کمک کدام ماده: پلی ساکارید تشکیل دهنده کپسول، پروتئین یا DNA؟
خیلی به حل این موضوع بستگی داشت، زیرا با ایجاد ماده ای که یک ویژگی ارثی را منتقل می کند - تشکیل یک کپسول، می توان پاسخ مطلوب را به دست آورد. با این حال، این کار برای مدتی طولانی امکان پذیر نبود. تنها 16 سال پس از آزمایشات F. Griffiths، در سال 1944، دانشمند آمریکایی A. Avery و همکارانش، با انجام یک سری آزمایشات واضح، توانستند با توجیه کامل ثابت کنند که پلی ساکارید و پروتئین هیچ ارتباطی با انتقال خواص ارثی پنوموکوک A-form.
در طی این آزمایشات، با کمک آنزیم خاصی، کپسول پلی ساکارید پنوموکوک های A شکل کشته شده حل شد و بررسی شد که آیا بقایای سلول A به انتقال اطلاعات ارثی به سلول های B شکل ادامه می دهد یا خیر. آنها ادامه می دهند. مشخص شد که پلی ساکارید به عنوان منبع اطلاعات ژنتیکی ناپدید می شود.
بنابراین، با روش حذف، مشخص شد که اطلاعات ارثی در سلول توسط مولکول DNA ذخیره و منتقل می شود. در واقع، هنگامی که DNA از بین رفت، تشکیل کپسول A از B غیر کپسوله شده متوقف شد.
پدیده تبدیل، یعنی تغییر ارثی در خواص یک شکل از باکتری ها تحت تأثیر موادی از شکل دیگر، تبدیل نامیده می شود. ماده ای که باعث دگرگونی می شود، عامل تبدیل نامیده می شود. مشخص شده است که آنها DNA هستند.
هر پروتئین با یک یا چند زنجیره پلی پپتیدی نشان داده می شود. بخشی از DNA که اطلاعات مربوط به یک زنجیره پلی پپتیدی را حمل می کند، ژن نامیده می شود. هر مولکول DNA حاوی ژن های مختلفی است. کل مولکول های DNA در یک سلول به عنوان حامل اطلاعات ژنتیکی عمل می کند. با توجه به ویژگی منحصر به فرد - توانایی تکثیر، که هیچ مولکول شناخته شده دیگری ندارد، DNA می تواند کپی شود. هنگام تقسیم، "نسخههای" DNA به دو سلول دختر واگرا میشوند که در نتیجه هر کدام از آنها همان اطلاعاتی را خواهند داشت که در سلول مادر موجود بود. از آنجایی که ژنها بخشهایی از مولکولهای DNA هستند، دو سلولی که در طول تقسیم تشکیل میشوند مجموعههای ژنهای یکسانی دارند. هر سلول یک ارگانیسم چند سلولی در طی تولید مثل جنسی از یک تخمک بارور شده در نتیجه تقسیمات متعدد بوجود می آید. این بدان معنی است که یک خطای تصادفی در ژن یک سلول در ژن میلیون ها نفر از فرزندان آن تکثیر می شود. به همین دلیل است که تمام گلبول های قرمز خون یک بیمار مبتلا به کم خونی داسی شکل، هموگلوبین آسیب دیده یکسانی دارند. این خطا در ژن حامل اطلاعات مربوط به زنجیره بتا پروتئین رخ داده است. یک کپی از یک ژن mRNA است. روی آن، مانند یک ماتریکس، پروتئین اشتباه هزاران بار در هر گلبول قرمز "چاپ" شده است. کودکان از طریق سلول های زایای خود ژن های فاسد را از والدین خود دریافت می کنند. اطلاعات ژنتیکی از یک سلول به سلول های دختر و از والدین به فرزندان منتقل می شود. ژن واحدی از اطلاعات ژنتیکی یا ارثی است.
جنین شناسی(از یونانی. جنین - جنین و لوگوس - کلمه، علم) - علم رشد جنین. دوره رشد فردی را مطالعه می کند که با لقاح شروع می شود و (در پستانداران و انسان) با تولد جنین پایان می یابد.
جنین شناسی تظاهرات مورفولوژیکی تشکیل از یک تخم بارور شده (زیگوت) یک ارگانیسم پیچیده چند سلولی را مطالعه می کند که شامل چهار بافت، چندین ده دیفرون سلولی، اندام ها، سیستم های اندام و دستگاه ها و بخش هایی از بدن است.
دانش جنین شناسی به درک بهتر آناتومی ماکروسکوپی کمک می کند. در یک سری منطقی با داده های جنین شناسی، حقایق تشریحی در مورد ساختار کلان و ریز بدن تأیید می کند که وابستگی های ساختاری در دوره رشد پس از تولد نتیجه طبیعی فرآیندهای ریخت زایی است که در جنین ها اتفاق می افتد.
جنین شناسی به شما امکان می دهد تظاهرات مورفولوژیکی، سودمندی اجرای برنامه ژنتیکی ذاتی در زیگوت و همچنین نقض این اجرا را تعیین کنید، که خود را در ناهنجاری های ساختاری و عملکردی رشد جنین نشان می دهد، که اغلب در بالینی یافت می شود. عملی است و تنها با دانستن جنین شناسی قابل توضیح است.
روند رشد جنین انسان، و همچنین سایر مهره داران، به چند مرحله تقسیم می شود که با ویژگی های کیفی و کمی مشخص می شود. مراحل اصلی جنین زایی عبارتند از: لقاح، له شدن، گاسترولاسیون، تشکیل بافت ها (هیستوژنز)، اندام ها (ارگانوژنز) و همچنین سیستم ها و دستگاه های اندام (سیستموژنز). برخی از کتابهای درسی و کتابهای راهنما نیز شامل پیشزایی، رشد سلولهای زایای زن و مرد، قبل از جنینزایی هستند.
این بخش عمدتاً به مراحل اولیه رشد جنین انسان می پردازد که با لقاح آغاز می شود و با تشکیل ابتدایی های بافتی و اندام های خارج جنینی پایان می یابد. در مورد بافت زایی، ارگانو و سیستم زایی، هنگام مطالعه ساختار اندام ها و سیستم های مربوطه در نظر گرفته می شوند.
لقاح تلفیقی از سلول های زایای زن و مرد است که منجر به تشکیل یک جنین تک سلولی می شود - یک زیگوت حاوی مجموعه دیپلوئیدی از کروموزوم ها و وراثت مادری و پدری را متحد می کند. لقاح در مجرای تخمک انجام می شود. مدت زمان این فرآیند حدود یک روز است.
زیگوت با میتوز شروع به تقسیم می کند. تقسیم آن را خرد کردن می نامند، زیرا در فرآیند تقسیم متوالی، زیگوت های سلول کاهش می یابد. در این رابطه، ابعاد کلی جنین چند سلولی در ابتدا ممکن است تقریباً با زیگوت ها یکسان باشد.
سلول های تشکیل شده در فرآیند خرد کردن، بلاستومر نامیده می شوند (از یونانی Blastos - جوانه، meros - قسمت). دوره شکاف با تقسیم زیگوت به دو بلاستومر آغاز می شود و تا تشکیل وزیکول (بلاستولا) ادامه می یابد. در انسان این دوره یک روز پس از لقاح شروع می شود و حدود 4-5 روز طول می کشد.
یکی از ویژگی های شکاف زیگوت پستانداران و انسان، ظاهر بلاستومرها است که از نظر اندازه و سرعت برش با هم تفاوت دارند. در ظاهر، بلاستومرهای کوچک و سبک (میکرومرها) قرار دارند که بیشتر تقسیم می شوند و تروفوبلاست یا میکروب زنده (از یونانی Tropho - I nourish) را تشکیل می دهند. در مرکز جنین، بلاستومرهای تیره بزرگ (ماکرومرها) با سیتوپلاسم بازوفیل وجود دارد، آنها کندتر تقسیم می شوند و با هم یک جنین یا ریشه جنین را تشکیل می دهند.
جنینی که حفره ندارد مورولا (از یونانی مورولا - توت) نامیده می شود. اگرچه این واژه مشخصه جنین های وتردار اولیه است (به عنوان مثال، نیزه، که در مراحل اولیه رشد در واقع شبیه توت است)، همچنین برای اشاره به جنین های اولیه مهره داران عالی که هنوز حفره ندارند، استفاده می شود. و تجمع متراکمی از سلول ها هستند.
سپس یک حفره کوچک پر از مایع در جنین ظاهر می شود و به شکل توپی به نام بلاستوسیست (بلاستولا) به خود می گیرد. دیواره بلاستوسیست توسط تروفوبلاست تشکیل می شود. جنین در سطح داخلی جنین قرار دارد.
در اواسط روز ششم، بیش از صد سلول در جنین وجود دارد. در این دوره، در رحم است، جایی که کاشته می شود (از لاتین Inplantatio - نفوذ)، یعنی عمیق شدن در غشای مخاطی. تعداد بلاستومرها در جنین افزایش می یابد.
در پایان هفته اول رشد، یک اپیتلیوم دو لایه از تروفوبلاست در حین لانه گزینی تشکیل می شود که لایه داخلی آن سیتوتروفوبلاست نامیده می شود. با سلول های مکعبی نشان داده می شود. در خارج یک پلاسمودیوتروفوبلاست وجود دارد که به آن سینسیتیوتروفوبلاست نیز می گویند، این یک سمپلاست است، یعنی. توده سیتوپلاسمی با هسته های متعدد به قلمروهای سلولی تقسیم نشده است.
نفوذ جنین به مخاط رحم با عمل تروفوبلاست همراه است که آنزیم های هیستولیتیک ترشح می کند که باعث تخریب موضعی (محلی) پوشش داخلی رحم - آندومتر می شود. جنین به ضخامت خود فرو می رود و آندومتر روی آن بسته می شود. بنابراین، رشد بیشتر جنین در مخاط رحم رخ می دهد.
تروفوبلاست در مراحل اولیه رشد جنین را تغذیه می کند، به طور فعال در لانه گزینی جنین و در تشکیل اندام موقت - کوریون شرکت می کند، که در تعامل با مخاط رحم، جفت را تشکیل می دهد. اندام جنینی که ارتباط بین جنین و بدن مادر را فراهم می کند. توسعه، ساختار و اهمیت عملکرد جفت در دوره های جنین شناسی و بافت شناسی مورد بحث قرار می گیرد.
جنین و سایر اندام های خارج جنینی از عناصر سلولی جنین رشد می کنند. یکی از ویژگی های اساسی رشد انسان، شکل گیری زودرس مزودرم خارج جنینی است. منبع وقوع آن جنین است. سلول ها از آن خارج می شوند، حفره بلاستوسیست را به شکل نوارها پر می کنند و تروفوبلاست را از داخل می پوشانند. مزودرم خارج جنینی به سرعت به مزانشیم خارج جنینی تبدیل می شود و در نتیجه پوسته بیرونی جنین از اپیتلیوم تروفوبلاستیک و مزانشیم تشکیل شده است. به آن کوریون می گویند. همانطور که در بالا ذکر شد، کوریون قسمت ژرمینال جفت را تشکیل می دهد، عضوی که ارتباط بین جنین و بدن مادر را فراهم می کند.
سپس گاسترولاسیون شروع می شود که به دو مرحله تقسیم می شود. در اول، لایه های جوانه تشکیل می شود، و در دوم - مجموعه محوری، که طرح کلی ساختار بدن مهره داران و انسان ها و پریموردیای ژرمینال را تعیین می کند، که از آن بافت های مختلف تشکیل می شود.
لایه های جوانه به لایه های اولیه سلولی گفته می شود که در مرحله اول گاسترولاسیون تشکیل شده اند. آنها از نظر مکان (برگ بیرونی - اکتودرم، داخلی - اندودرم و میانی - مزودرم)، اندازه و ساختار سلول ها و جهت رشد بیشتر متفاوت هستند. مجموعه محوری شامل لوله عصبی، وتر (رشته پشتی)، مزودرم پشتی، واقع در دو طرف لوله عصبی و وتر، و روده اولیه است. در مرحله دوم گاسترولاسیون، پایه های جنینی نیز تشکیل می شوند - مجتمع های سلولی که از لایه های جوانه ایجاد می شوند و بافت های مختلف را شروع می کنند. بنابراین، بافت های مختلف از یک لایه جنینی تشکیل می شود.
تظاهرات مورفولوژیکی فاز اول گاسترولاسیون، که در یک جنین 7.5 هفته ای مشاهده می شود، تقسیم جنین به دو لایه - خارجی (اپی بلاست) و داخلی (هیپوبلاست) است.
اپی بلاست دارای اجزای اکتودرم، مزودرم ژرمینال، وتر (رشته پشتی) و آندودرم ژرمینال (روده) است. هیپوبلاست پایه ای از اندودرم خارج جنینی یا زرده است که بخشی از لایه جوانه داخلی است که متعاقباً بخشی از یکی از اندام های موقت - کیسه زرده - می شود.
سلول های اندودرم جدا شده از اپی بلاست در اطراف حفره تشکیل شده در نواحی پایینی مزودرم خارج جنینی شروع به رشد می کنند. به تدریج شکل کاسه ای به خود می گیرند که لبه های آن با هم رشد می کنند و در نتیجه وزیکول زرده به وجود می آید. این فرآیند با شروع تشکیل کیسه زرده شبیه به آنچه که مشخصه رشد این اندام خارج جنینی در پستانداران است.
متعاقبا، پس از شروع کاشت، یک حفره آمنیوتیک اولیه در اپی بلاست به دلیل ظاهر شدن مایعی که سلول ها را از هم جدا می کند ظاهر می شود. سپس سقف این حفره (وزیکول آمنیوتیک) باز می شود و به طور موقت توسط بخشی از سیتوتروفوبلاست از خارج خارج می شود. پس از آن، دیواره های وزیکول آمنیوتیک به سمت بالا رشد می کنند و به هم می پیوندند، در نتیجه حفره وزیکول آمنیوتیک دوباره بسته می شود و توسط سلول های اپی بلاست احاطه می شود. این فرآیند شبیه به ظاهر چین های آمنیوتیک در پرندگان و پستانداران است.
از 9 تا 14 روز رشد، جنین انسان دارای ساختار زیر است: دیواره بیرونی تشکیل شده توسط کوریون از مزانشیم خارج جنینی و اپیتلیوم تروفوبلاستیک تشکیل شده است. دومی، همانطور که قبلا ذکر شد، شامل سیتوتروفوبلاست و پلاسمودیوتروفوبلاست است. در دیواره ضخیم جنین، که به عمق دیواره رحم تبدیل شده است، مزانشیم خارج جنینی حاوی دو وزیکول است که با یکدیگر در تماس هستند - آمنیوتیک و زرده. ناحیه وزیکول آمنیوتیک در مجاورت وزیکول ویتلین یک ضخیم شدن ایجاد می کند - به اصطلاح سپر ژرمینال که بدن جنین از آن تشکیل می شود. از مزانشیم، دو وزیکول محدود می شود، نواری از سلول ها به کوریون کشیده می شود - پای آمنیوتیک، محل تخلیه آن مربوط به انتهای خلفی بدن جنین است.
به لطف فرآیندهایی که در بالا توضیح داده شد، جنین دارای سه اندام خارج جنینی است - کوریون که در تشکیل جفت، آمنیون و کیسه زرده نقش دارد. بیشتر وزیکول آمنیوتیک یک اندام خارج جنینی را تشکیل می دهد - آمنیون که یکی از وظایف آن تشکیل مایع آمنیوتیک است - یک محیط آبی مصنوعی برای جنین در حال رشد. از هیپوبلاست، یک وزیکول زرده (کیسه) تشکیل می شود که در انسان و اکثر پستانداران در زرده وجود ندارد، اما نقش مهمی را به عنوان اولین اندام خونساز درگیر در توسعه سلول های زایای اولیه ایفا می کند.
بنابراین، در فرآیند رشد جنینی انسان، ظهور تعدادی از اندام های خارج جنینی (کوریون، آمنیون و کیسه زرده) مقدم بر پیدایش بدن جنین است که دلیلی بر ایجاد اولیه شرایط لازم است. برای رشد جنین
در روز پانزدهم رشد داخل رحمی، مرحله دوم گاسترولاسیون شروع می شود که هم در پرندگان و هم در پستانداران جفتی به یک شکل اتفاق می افتد. انتقال از مرحله اول گاسترولاسیون به مرحله دوم به تدریج انجام می شود و علائم اول (تشکیل لایه های جوانه) و دوم (تشکیل مجموعه ای از اندام های محوری) را با هم ترکیب می کند. علامت خارجی فاز دوم گاسترولاسیون، ظاهر شدن یک نوار دراز سلولی در انتهای خلفی سپر ژرمینال است که به آن نوار اولیه می گویند. در انتهای قدامی آن، یک برآمدگی جزئی از سلول ها تشکیل می شود - ندول اولیه یا هنسن.
در بالای گره اولیه، یک فرورفتگی کوچک ظاهر می شود - حفره اولیه. در قسمت مرکزی نوار اولیه، یک شیار اولیه ظاهر می شود که ادامه حفره اولیه است. بخشی از ماده لایه بیرونی زاینده از طریق حفره اولیه به سمت داخل فرو میرود و بخشی از آندودرم قدامی است و به اصطلاح صفحه پیشکوردال را تشکیل میدهد که از آن کانال گوارشی قدامی با پوشش اپیتلیال و دو جفت سومیت قدامی (بخشهای تکهشده پشتی) تشکیل میشود. مزودرم) تشکیل می شوند.
جهت رگه اولیه مکان اندام های محوری و در نتیجه طرح کلی ساختار بدن مهره داران را تعیین می کند.
بخشی از سلول های اپی بلاست از طریق رگه اولیه مهاجرت می کند و با نفوذ به قسمت زیرین هیپوبلاست، پایه اندودرم روده را تشکیل می دهد که متعاقباً در رشد روده شرکت می کند. قسمت دوم هیپوبلاست، همانطور که در بالا ذکر شد، به تشکیل اندودرم زرده - لایه داخلی کیسه زرده می رود.
در نتیجه مهاجرت سلولی از اپی بلاست از طریق رگه اولیه، مزودرم ژرمینال نیز بوجود می آید که در جهات جانبی زیر اپی بلاست گسترش می یابد.
منشأ رشته پشتی - وتر - ارتباط نزدیکی با تشکیل مزودرم ژرمینال دارد. پایه آن به شکل فرآیند به اصطلاح آکوردال (یا اصلی) تشکیل می شود - رشته ای از سلول ها که به دلیل مزودرم ژرمینال تشکیل شده است که از گره اصلی (هنزن) بین اپی بلاست و هیپوبلاست رشد می کند. نوتوکورد محل ستون فقرات آینده را تعیین می کند، زیرا مهره ها در اطراف آن تشکیل می شوند و به طور قابل توجهی بر رشد سیستم عصبی تأثیر می گذارد. در پستانداران بالاتر، بقایای نوتوکورد در دوره رشد پس از تولد به عنوان بخشی از هسته پالپوزوس، هسته پالپوزوس و دیسک های بین مهره ای حفظ می شود.
لایه بالایی سلول های محافظ ژرمینال پس از تشکیل نوتوکورد به اکتودرم تبدیل می شود. تحت تأثیر وتر یا کوردومزودرم بر روی اکتودرم موجود در بالای آن، دومی ضخیم می شود و به صفحه عصبی تبدیل می شود، سپس به یک پیچ عصبی دراز و بعداً به لوله عصبی تبدیل می شود که به دلیل آن سیستم عصبی مرکزی و محیطی شکل گرفت.
در پایان دوم - آغاز هفته سوم رشد جنینی، چهارمین اندام خارج جنینی، آلانتویس، گذاشته می شود. در ابتدا، به نظر می رسد بیرون آمدن کیسه زرده، کورکورانه به پایان می رسد و به ساقه آمنیوتیک رشد می کند. بعداً (پس از تشکیل روده)، آلانتویس از دیواره قدامی لوله روده خلفی خارج می شود. به دنبال تشکیل لوله عصبی، لایه های مزودرم در کنار آن و نوتوکورد در معرض تقسیم بندی قرار می گیرند. بخشهای جانبی مزودرم که بعداً بخشی از قسمت شکمی جنین میشوند و به همین دلیل مزودرم شکمی نامیده میشوند، به دو لایه تقسیم میشوند: لایه بیرونی در تماس با اکتودرم (لایه جداری) و لایه داخلی، متصل میشود. به اندودرم (لایه احشایی). سومیت ها توسط رشته های نازکی از سلول ها به مزودرم شکمی به نام مزودرم میانی متصل می شوند. سپس سومیت ها به سه قسمت پشتی جانبی (درماتوم)، مدیوونترال (اسکلروتوم) و میانی (میوتوم) تقسیم می شوند. با ظهور سومیت ها، دوره سومیتی رشد جنین آغاز می شود. در روز 21، 2-3 جفت سومیت در ترکیب آن یافت می شود. در روز 23 - 10 جفت؛ در روز 25 - 14 جفت؛ در روز 27 - 25 جفت؛ در پایان هفته 5 - 43-44 جفت.
شکاف های بین لایه های جوانه و اندام های اصلی با بافت همبند ژرمینال پر می شود - مزانشیم که از سلول های دوکی یا سطلی شکل تشکیل شده است که با فرآیندها در تماس هستند و در یک ماده بین سلولی بی شکل غوطه ور می شوند. مزانشیم عمدتاً از مزودرم تشکیل می شود، اما سایر لایه های جوانه، که اکتودرم از آنها نیز در توسعه آن شرکت می کنند (M. F. Kashchenko).
هنگامی که لایه احشایی مزودرم شکمی با آندودرم روده تماس پیدا می کند، بدن جنین شروع به جدا شدن از اندام های خارج جنینی توسط چین های تنه می کند. این فرآیند با انحراف شکمی لبه های جنین شروع می شود که در نتیجه تحدب پشتی به دست می آید. سپس لبه های منحنی جنین در قسمت های اصلی و دمی شروع به عمیق شدن می کنند. گسترش این چین های جداکننده منجر به تشکیل دیواره های جانبی و شکمی جنین و همچنین به وجود آمدن ساقه به اصطلاح زرده می شود که آن را به کیسه زرده متصل می کند. با عمیق شدن چین تنه، آندودرم روده و ورقه احشایی مزودرم شکمی به یک لوله تبدیل می شود که منجر به تشکیل روده می شود. در حالی که دومی شامل اندودرم روده است، کیسه زرده حاوی اندودرم زرده است. آلانتویس که قبلاً ظاهر برآمدگی کیسه زرده داشت، اکنون از روده عقبی گسترش یافته است.
با رشد غشای آمنیوتیک، حجم آمنیون افزایش می یابد که به کوریون نزدیک شده و با آن جوش می خورد. در همان زمان، پای آمنیوتیک از ناحیه دمی به سمت دیواره های شکمی جنین حرکت می کند و با آمنیون متصل می شود و بند ناف (بند ناف) را تشکیل می دهد. آلانتوا که قبلاً به ساقه آمنیوتیک نفوذ کرده بود تا حدی در بند ناف قرار می گیرد. در فرآیند این دگرگونی ها کیسه زرده از جنین خارج می شود و به دلیل مجرای زرده-روده ای لوله ای، ارتباط آن با روده حفظ می شود. دومی همراه با آلانتویس نیز بخشی از بند ناف است. متعاقباً آلانتویس و مجرای زرده-روده ای که بخشی از بند ناف هستند کاهش می یابد.
 مراحل جنین زایی I - لقاح و تشکیل زیگوت. II - خرد شدن و تشکیل بلاستولا (بلاستوسیست)؛ III - گاسترولاسیون - تشکیل لایه های جوانه و مجموعه ای از اندام های محوری. IV - هیستوژنز و اندام زایی اندام های زایایی و خارج جنینی. V - سیستم زایی.
مراحل جنین زایی I - لقاح و تشکیل زیگوت. II - خرد شدن و تشکیل بلاستولا (بلاستوسیست)؛ III - گاسترولاسیون - تشکیل لایه های جوانه و مجموعه ای از اندام های محوری. IV - هیستوژنز و اندام زایی اندام های زایایی و خارج جنینی. V - سیستم زایی.
 دوره قبل از تولد از لحظه لقاح و تشکیل زیگوت شروع می شود و 280 روز طول می کشد. جنین زایی به دو دسته تقسیم می شود: - دوره اولیه (هفته اول) - دوره جنینی (از هفته دوم تا نهم) - دوره جنینی (از 8-9 هفته تا تولد).
دوره قبل از تولد از لحظه لقاح و تشکیل زیگوت شروع می شود و 280 روز طول می کشد. جنین زایی به دو دسته تقسیم می شود: - دوره اولیه (هفته اول) - دوره جنینی (از هفته دوم تا نهم) - دوره جنینی (از 8-9 هفته تا تولد).

 GAMETOGENESIS ✓ گنوسیت های سلول های زایای اولیه (گنوبلاست ها) برای اولین بار در دیواره کیسه زرده (در پایان هفته سوم زندگی جنین انسان) شناسایی می شوند. ✓ گنوسیت ها سلول های بی تفاوت هستند: آنها به سلول های نر و ماده تقسیم نمی شوند. ✓ گنوسیت ها به ناحیه تناسلی غدد جنسی مهاجرت می کنند و به اسپرماتوگونی (در بیضه) یا اووگونیا (در تخمدان) تمایز می یابند. این فرآیند در هفته ششم زندگی جنین آغاز می شود.
GAMETOGENESIS ✓ گنوسیت های سلول های زایای اولیه (گنوبلاست ها) برای اولین بار در دیواره کیسه زرده (در پایان هفته سوم زندگی جنین انسان) شناسایی می شوند. ✓ گنوسیت ها سلول های بی تفاوت هستند: آنها به سلول های نر و ماده تقسیم نمی شوند. ✓ گنوسیت ها به ناحیه تناسلی غدد جنسی مهاجرت می کنند و به اسپرماتوگونی (در بیضه) یا اووگونیا (در تخمدان) تمایز می یابند. این فرآیند در هفته ششم زندگی جنین آغاز می شود.
 اسپرماتوژنز در لوله های پیچ خورده بیضه رخ می دهد. در فرآیند اسپرم زایی، 4 مرحله (دوره) متمایز می شود: تولید مثل (تکثیر) تشکیل بلوغ رشد (اسپرمیوژنز).
اسپرماتوژنز در لوله های پیچ خورده بیضه رخ می دهد. در فرآیند اسپرم زایی، 4 مرحله (دوره) متمایز می شود: تولید مثل (تکثیر) تشکیل بلوغ رشد (اسپرمیوژنز).
 I. مرحله تولید مثل: سلول های تولید مثلی نر توسط سلول های دیپلوئیدی - SPERMATOGONIA که به صورت MITOTICally تقسیم می شوند نشان داده می شوند. 3 نوع اسپرماتوگونی وجود دارد (شکل 1): تیره و اسپرماتوگونیا سلول های بنیادی واقعی هستند که در برابر عوامل مضر مقاوم هستند و به ندرت تقسیم می شوند. نور اسپرماتوگونی A سلول های نیمه بنیادی هستند که قادر به تقسیمات میتوزی مکرر هستند (نتیجه تقسیم: دو اسپرماتوگونی A تشکیل می شود یا یک A و یک B-). نور در اسپرماتوگونی، سلول های متعهد برای میوز آماده می شوند و وارد فاز رشد می شوند.
I. مرحله تولید مثل: سلول های تولید مثلی نر توسط سلول های دیپلوئیدی - SPERMATOGONIA که به صورت MITOTICally تقسیم می شوند نشان داده می شوند. 3 نوع اسپرماتوگونی وجود دارد (شکل 1): تیره و اسپرماتوگونیا سلول های بنیادی واقعی هستند که در برابر عوامل مضر مقاوم هستند و به ندرت تقسیم می شوند. نور اسپرماتوگونی A سلول های نیمه بنیادی هستند که قادر به تقسیمات میتوزی مکرر هستند (نتیجه تقسیم: دو اسپرماتوگونی A تشکیل می شود یا یک A و یک B-). نور در اسپرماتوگونی، سلول های متعهد برای میوز آماده می شوند و وارد فاز رشد می شوند.
 بخش دیواره لوله پیچ خورده بیضه 1 - سلول های میوئید دیوار. 2 - غشای پایه؛ 3 - اسپرماتوگونی A; 4 - اسپرماتوگونی B; 5 - اسپرماتوسیت درجه اول; 6 - اسپرماتوسیت درجه دوم؛ 7 - اسپرماتیدها در مراحل اولیه رشد. 8 - اسپرماتیدها در پایان نمو. 9 - سلول سرتولی.
بخش دیواره لوله پیچ خورده بیضه 1 - سلول های میوئید دیوار. 2 - غشای پایه؛ 3 - اسپرماتوگونی A; 4 - اسپرماتوگونی B; 5 - اسپرماتوسیت درجه اول; 6 - اسپرماتوسیت درجه دوم؛ 7 - اسپرماتیدها در مراحل اولیه رشد. 8 - اسپرماتیدها در پایان نمو. 9 - سلول سرتولی.
 II. فاز رشد: در اسپرماتوگونی حجم آنها (4 برابر) افزایش می یابد و وارد پروفاز اولین تقسیم میوز می شوند و از لایه بازال به سمت لومن توبول حرکت می کنند - اسپرماتوسیت های درجه یک (دیپلوئید).
II. فاز رشد: در اسپرماتوگونی حجم آنها (4 برابر) افزایش می یابد و وارد پروفاز اولین تقسیم میوز می شوند و از لایه بازال به سمت لومن توبول حرکت می کنند - اسپرماتوسیت های درجه یک (دیپلوئید).
 III. مرحله بلوغ: در نتیجه تقسیم 1 میوز (کاهش)، SPERMATOCYTE های مرتبه 2 تشکیل می شوند که به سرعت وارد تقسیم 2 میوز (معادله) می شوند و سلول های حاصل را SPERMATIDS (هاپلوئید) می نامند. با شروع اسپرماتوگونی نوع B، سلول های دختر در طول تقسیم به طور کامل از یکدیگر جدا نمی شوند، اما با پل های سیتوپلاسمی به هم متصل می شوند و سینسیتیوم را تشکیل می دهند. فقط اسپرم های بالغ از سنسیتیوم جدا می شوند.
III. مرحله بلوغ: در نتیجه تقسیم 1 میوز (کاهش)، SPERMATOCYTE های مرتبه 2 تشکیل می شوند که به سرعت وارد تقسیم 2 میوز (معادله) می شوند و سلول های حاصل را SPERMATIDS (هاپلوئید) می نامند. با شروع اسپرماتوگونی نوع B، سلول های دختر در طول تقسیم به طور کامل از یکدیگر جدا نمی شوند، اما با پل های سیتوپلاسمی به هم متصل می شوند و سینسیتیوم را تشکیل می دهند. فقط اسپرم های بالغ از سنسیتیوم جدا می شوند.

 IV. مرحله تشکیل (اسپرمیوژنز): حدود 50 روز در طول تشکیل اسپرم بالغ از اسپرماتیدها، موارد زیر رخ می دهد: تراکم کروماتین هسته ای. تراکم کروماتین منجر به تشکیل یک هسته فوق فشرده می شود که تحرک اسپرم را افزایش می دهد و از ژنوم در برابر آسیب محافظت می کند. تشکیل آکروزوم از عناصر مجموعه گلژی؛ آکروزوم - نوع خاصی از لیزوزوم، حاوی آنزیم های هیدرولیتیک (مانند هیالورونیداز و غیره) برای از بین بردن غشای تخم مرغ. سانتریول ها به سمت قطب مخالف هسته حرکت می کنند. یک رشته محوری (آکسونم) تاژک از سانتریول دیستال تشکیل می شود. میتوکندری به شکل یک قسمت مارپیچی آکسونم را احاطه کرده و یک ناحیه ضخیم شده - یک بخش میانی را تشکیل می دهد. میتوکندری انرژی برای حرکت اسپرم فراهم می کند. سیتوپلاسم به حداقل کاهش می یابد. پس از ریزش سیتوپلاسم اضافی، اسپرم از انجمن سلولی مشترک - سینسیتیوم - جدا شده و آزاد می شود.
IV. مرحله تشکیل (اسپرمیوژنز): حدود 50 روز در طول تشکیل اسپرم بالغ از اسپرماتیدها، موارد زیر رخ می دهد: تراکم کروماتین هسته ای. تراکم کروماتین منجر به تشکیل یک هسته فوق فشرده می شود که تحرک اسپرم را افزایش می دهد و از ژنوم در برابر آسیب محافظت می کند. تشکیل آکروزوم از عناصر مجموعه گلژی؛ آکروزوم - نوع خاصی از لیزوزوم، حاوی آنزیم های هیدرولیتیک (مانند هیالورونیداز و غیره) برای از بین بردن غشای تخم مرغ. سانتریول ها به سمت قطب مخالف هسته حرکت می کنند. یک رشته محوری (آکسونم) تاژک از سانتریول دیستال تشکیل می شود. میتوکندری به شکل یک قسمت مارپیچی آکسونم را احاطه کرده و یک ناحیه ضخیم شده - یک بخش میانی را تشکیل می دهد. میتوکندری انرژی برای حرکت اسپرم فراهم می کند. سیتوپلاسم به حداقل کاهش می یابد. پس از ریزش سیتوپلاسم اضافی، اسپرم از انجمن سلولی مشترک - سینسیتیوم - جدا شده و آزاد می شود.


 I. مرحله تولید مثل: بر خلاف اسپرماتوژنز، مرحله تولید مثل در اووژنز منحصراً در طول دوره رشد جنینی انجام می شود. اووگونیا با میتوز تقسیم می شود. بیشتر سلول های دختر حاصل می میرند (این فرآیند آترزی نامیده می شود)، برخی از آنها وارد فاز رشد می شوند (در پروفاز اولین تقسیم میوز). تقسیم میتوزی اووگون ها در ماه هفتم جنین زایی متوقف می شود. در بدو تولد، تمام تخمکهای تخمدان، اووسیتهای مرتبه اول (اووسیت اولیه) هستند.
I. مرحله تولید مثل: بر خلاف اسپرماتوژنز، مرحله تولید مثل در اووژنز منحصراً در طول دوره رشد جنینی انجام می شود. اووگونیا با میتوز تقسیم می شود. بیشتر سلول های دختر حاصل می میرند (این فرآیند آترزی نامیده می شود)، برخی از آنها وارد فاز رشد می شوند (در پروفاز اولین تقسیم میوز). تقسیم میتوزی اووگون ها در ماه هفتم جنین زایی متوقف می شود. در بدو تولد، تمام تخمکهای تخمدان، اووسیتهای مرتبه اول (اووسیت اولیه) هستند.
 II. مرحله رشد: مرحله رشد مربوط به عمر تخمک اولیه است که در پروفاز تقسیم اول میوز قرار دارد به دو دوره رشد کوچک (قبل از بلوغ) و رشد بزرگ تقسیم می شود. تا زمان تولد، حدود 2 میلیون تخمک اولیه در تخمدان وجود دارد. در سن بلوغ، اکثر آنها می میرند و حدود 400000 تخمک باقی می مانند. رشد تخمک در فولیکول ها اتفاق می افتد، ساختارهایی که در آن تخمک توسط سلول های فولیکولی اپیتلیال احاطه شده است. در طول دوره رشد کوچک، دو نوع فولیکول وجود دارد: فولیکول های اولیه (یک تخمک توسط یک لایه منفرد از سلول های فولیکولی مسطح احاطه شده است) و فولیکول های اولیه (یک براق (پوسته شفاف) ظاهر می شود، سلول های فولیکولی مکعبی یا منشوری هستند). ورود تخمک ها به دوره رشد زیاد فقط تحت تأثیر هورمون ها (هورمون محرک فولیکول) اتفاق می افتد. این دوره از اووژنز مربوط به فولیکول های در حال رشد (ثانویه و سوم) است. دوره رشد می تواند از 12 تا 50 سال طول بکشد.
II. مرحله رشد: مرحله رشد مربوط به عمر تخمک اولیه است که در پروفاز تقسیم اول میوز قرار دارد به دو دوره رشد کوچک (قبل از بلوغ) و رشد بزرگ تقسیم می شود. تا زمان تولد، حدود 2 میلیون تخمک اولیه در تخمدان وجود دارد. در سن بلوغ، اکثر آنها می میرند و حدود 400000 تخمک باقی می مانند. رشد تخمک در فولیکول ها اتفاق می افتد، ساختارهایی که در آن تخمک توسط سلول های فولیکولی اپیتلیال احاطه شده است. در طول دوره رشد کوچک، دو نوع فولیکول وجود دارد: فولیکول های اولیه (یک تخمک توسط یک لایه منفرد از سلول های فولیکولی مسطح احاطه شده است) و فولیکول های اولیه (یک براق (پوسته شفاف) ظاهر می شود، سلول های فولیکولی مکعبی یا منشوری هستند). ورود تخمک ها به دوره رشد زیاد فقط تحت تأثیر هورمون ها (هورمون محرک فولیکول) اتفاق می افتد. این دوره از اووژنز مربوط به فولیکول های در حال رشد (ثانویه و سوم) است. دوره رشد می تواند از 12 تا 50 سال طول بکشد.
 III. مرحله بلوغ اولین تقسیم میوز درست قبل از تخمک گذاری کامل می شود (تخمک گذاری عبارت است از پارگی فولیکول سوم و آزاد شدن تخمک از تخمدان به داخل حفره شکمی). در نتیجه میوز I، یک تخمک مرتبه دوم (تخمک ثانویه) تشکیل می شود - یک سلول هاپلوئید. تخمک مرتبه دوم وارد تقسیم دوم میوز می شود، اما این تقسیم در مرحله متافاز مسدود شده و تنها در شرایط لقاح کامل می شود. در یک چرخه قاعدگی-تخمدانی (28 روز)، معمولا یک تخمک تخمک گذاری می شود. بنابراین، در طول دوره باروری زندگی یک زن (~ 3040 سال)، تنها حدود 450 تخمک به بلوغ می رسد - آمادگی برای لقاح.
III. مرحله بلوغ اولین تقسیم میوز درست قبل از تخمک گذاری کامل می شود (تخمک گذاری عبارت است از پارگی فولیکول سوم و آزاد شدن تخمک از تخمدان به داخل حفره شکمی). در نتیجه میوز I، یک تخمک مرتبه دوم (تخمک ثانویه) تشکیل می شود - یک سلول هاپلوئید. تخمک مرتبه دوم وارد تقسیم دوم میوز می شود، اما این تقسیم در مرحله متافاز مسدود شده و تنها در شرایط لقاح کامل می شود. در یک چرخه قاعدگی-تخمدانی (28 روز)، معمولا یک تخمک تخمک گذاری می شود. بنابراین، در طول دوره باروری زندگی یک زن (~ 3040 سال)، تنها حدود 450 تخمک به بلوغ می رسد - آمادگی برای لقاح.
 تقسیم میوز در تخمزایی از این جهت متفاوت است که با سیتوتومی نابرابر پیش میرود: در میوز I، کروموزومها به طور مساوی بین سلولهای دختر توزیع میشوند، اما تنها یکی از سلولها (اووسیت ثانویه) تقریباً کل سیتوپلاسم را حفظ میکند. سلول دوم (اولین جسم جهت دار قطبی یا کاهشی) دارای اندازه بسیار کوچک، هسته کوچک و حداقل سیتوپلاسم است.
تقسیم میوز در تخمزایی از این جهت متفاوت است که با سیتوتومی نابرابر پیش میرود: در میوز I، کروموزومها به طور مساوی بین سلولهای دختر توزیع میشوند، اما تنها یکی از سلولها (اووسیت ثانویه) تقریباً کل سیتوپلاسم را حفظ میکند. سلول دوم (اولین جسم جهت دار قطبی یا کاهشی) دارای اندازه بسیار کوچک، هسته کوچک و حداقل سیتوپلاسم است.


 تخم انسان در درجه دوم اولیگولسیتال و ایزوولسیتال است: مقدار نسبتا کمی زرده به طور مساوی در سیتوپلاسم توزیع شده است. علاوه بر این، در تکامل، این امر ثانویه است: برای اولین بار این نوع تخم مرغ در نیزه یافت می شود.
تخم انسان در درجه دوم اولیگولسیتال و ایزوولسیتال است: مقدار نسبتا کمی زرده به طور مساوی در سیتوپلاسم توزیع شده است. علاوه بر این، در تکامل، این امر ثانویه است: برای اولین بار این نوع تخم مرغ در نیزه یافت می شود.
 ساختارهای خاص سیتوپلاسم تخم گرانول های زرد - گرانول های غشایی حاوی فسفو و لیپوپروتئین ها - فسفوویتین و لیپوویتلین هستند. برخی از این مواد در کبد یک زن تشکیل می شوند، برخی دیگر مستقیماً در تخمک I. گرانول های قشری - این گرانول ها با قرار گرفتن در زیر پلاسمالما حاوی آنزیم هایی هستند که پس از لقاح، در واکنش قشری شرکت می کنند.
ساختارهای خاص سیتوپلاسم تخم گرانول های زرد - گرانول های غشایی حاوی فسفو و لیپوپروتئین ها - فسفوویتین و لیپوویتلین هستند. برخی از این مواد در کبد یک زن تشکیل می شوند، برخی دیگر مستقیماً در تخمک I. گرانول های قشری - این گرانول ها با قرار گرفتن در زیر پلاسمالما حاوی آنزیم هایی هستند که پس از لقاح، در واکنش قشری شرکت می کنند.
 ویژگی های ترکیب سیتوپلاسم تخم مرغ در سیتوپلاسم - محتوای بسیار بالایی از اجزای سیستم سنتز پروتئین (ریبوزوم ها، m. RNA، m. RNA). ✓هیچ سانتریول وجود ندارد - از این نظر، توانایی تقسیم تنها زمانی بازسازی می شود که سانتریول های اسپرم وارد سلول شوند. ✓ میکروویلی در سطح پلاسمالما وجود دارد. ✓ تخمک (به طور دقیق تر، تخمک II)، مانند سلول های قبل از آن، توسط پوسته هایی احاطه شده است: براق یا شفاف (zona pellucida یا Zp) و دانه ای که توسط سلول های فولیکولی تشکیل شده است.
ویژگی های ترکیب سیتوپلاسم تخم مرغ در سیتوپلاسم - محتوای بسیار بالایی از اجزای سیستم سنتز پروتئین (ریبوزوم ها، m. RNA، m. RNA). ✓هیچ سانتریول وجود ندارد - از این نظر، توانایی تقسیم تنها زمانی بازسازی می شود که سانتریول های اسپرم وارد سلول شوند. ✓ میکروویلی در سطح پلاسمالما وجود دارد. ✓ تخمک (به طور دقیق تر، تخمک II)، مانند سلول های قبل از آن، توسط پوسته هایی احاطه شده است: براق یا شفاف (zona pellucida یا Zp) و دانه ای که توسط سلول های فولیکولی تشکیل شده است.
 زونا پلوسیدا زونا پلوسیدا از انواع مختلفی از گلیکوپروتئین ها (Zp 1، Zp 2، Zp 3) و گلیکوزامینوگلیکان ها تشکیل شده است. گلیکوپروتئین های کسر Zp 3 گیرنده های اسپرم هستند و گلیکوپروتئین های کسر Zp 2 از پلی اسپرمی بعد از واکنش قشر مغز جلوگیری می کنند. اجزای زونا پلوسیدا توسط سلول های فولیکولی سنتز می شوند. غلاف دانه ای: سلول های فولیکولی غلاف دانه ای بخشی از اپیتلیوم فولیکولی هستند و فرآیندهای طولانی در زونا پلوسیدا نفوذ می کنند. تاج تابشی: منظور از تاج تابشی فقط قسمتهای داخلی سلولهای فولیکولی است که فرآیندهایی از آنها امتداد مییابد، یا کل لایه دانهای و زونا پلوسیدا. اجسام کاهشی: در جایی در غشاها یا زیر آنها اجسام کاهشی وجود دارد - محصولات دیگر (به جز تخم مرغ) از دو بخش میوز.
زونا پلوسیدا زونا پلوسیدا از انواع مختلفی از گلیکوپروتئین ها (Zp 1، Zp 2، Zp 3) و گلیکوزامینوگلیکان ها تشکیل شده است. گلیکوپروتئین های کسر Zp 3 گیرنده های اسپرم هستند و گلیکوپروتئین های کسر Zp 2 از پلی اسپرمی بعد از واکنش قشر مغز جلوگیری می کنند. اجزای زونا پلوسیدا توسط سلول های فولیکولی سنتز می شوند. غلاف دانه ای: سلول های فولیکولی غلاف دانه ای بخشی از اپیتلیوم فولیکولی هستند و فرآیندهای طولانی در زونا پلوسیدا نفوذ می کنند. تاج تابشی: منظور از تاج تابشی فقط قسمتهای داخلی سلولهای فولیکولی است که فرآیندهایی از آنها امتداد مییابد، یا کل لایه دانهای و زونا پلوسیدا. اجسام کاهشی: در جایی در غشاها یا زیر آنها اجسام کاهشی وجود دارد - محصولات دیگر (به جز تخم مرغ) از دو بخش میوز.


 غشای پلاسمایی سر: حاوی پروتئینهای ویژهای است که در تاکسیهای اسپرمها و اتصال به تخمک نقش دارند. آکروزوم: آکروزوم یک کیسه غشایی مسطح است که هسته را دو درپوش دارد. حاوی آنزیم های لیتیک (آکروزین، هیالورونیداز و غیره) است که غشاهای تخم را از بین می برد. هسته: به شدت فشرده شده و حاوی مجموعه ای از کروموزوم هاپلوئید است.
غشای پلاسمایی سر: حاوی پروتئینهای ویژهای است که در تاکسیهای اسپرمها و اتصال به تخمک نقش دارند. آکروزوم: آکروزوم یک کیسه غشایی مسطح است که هسته را دو درپوش دارد. حاوی آنزیم های لیتیک (آکروزین، هیالورونیداز و غیره) است که غشاهای تخم را از بین می برد. هسته: به شدت فشرده شده و حاوی مجموعه ای از کروموزوم هاپلوئید است.
 گردن دم یا بخش اتصال: شامل 2 سانتریول. از یکی از آنها آکسونم یا رشته محوری دم شروع می شود که طبق طرح (9 x2) + 2 توسط میکروتوبول ها تشکیل شده است. غشا و پلاسمالما قسمت اصلی یا اصلی: در اینجا، در اطراف آکسونم، یک غلاف فیبریلار (9 فیبریل بیرونی و یک غلاف فیبری) و همچنین پلاسمولما وجود دارد. قسمت پایانی: در این قسمت فقط پلاسمولما در اطراف آکسونم باقی می ماند.
گردن دم یا بخش اتصال: شامل 2 سانتریول. از یکی از آنها آکسونم یا رشته محوری دم شروع می شود که طبق طرح (9 x2) + 2 توسط میکروتوبول ها تشکیل شده است. غشا و پلاسمالما قسمت اصلی یا اصلی: در اینجا، در اطراف آکسونم، یک غلاف فیبریلار (9 فیبریل بیرونی و یک غلاف فیبری) و همچنین پلاسمولما وجود دارد. قسمت پایانی: در این قسمت فقط پلاسمولما در اطراف آکسونم باقی می ماند.
 انزال نر دارای حجم 2-3 میلی لیتر و دارای 200-350 میلیون اسپرم است و در انسان سر اسپرم به شدت پهن است.
انزال نر دارای حجم 2-3 میلی لیتر و دارای 200-350 میلیون اسپرم است و در انسان سر اسپرم به شدت پهن است.
 ترکیب سلولی انزال انسان A- بالغ، B- نابالغ 1، 2 - اسپرم معمولی 3-12 - اسپرم آتیپیک 6-7 - ناهنجاری در شکل سر و آکروزوم 8-9 - ناهنجاری تاژک 13-18 - نابالغ سلول های زایای مردانه 19 - سلول های اپیتلیال 20 - 22 - لکوسیت ها
ترکیب سلولی انزال انسان A- بالغ، B- نابالغ 1، 2 - اسپرم معمولی 3-12 - اسپرم آتیپیک 6-7 - ناهنجاری در شکل سر و آکروزوم 8-9 - ناهنجاری تاژک 13-18 - نابالغ سلول های زایای مردانه 19 - سلول های اپیتلیال 20 - 22 - لکوسیت ها
 اسپرمها (بر خلاف تخممرغ) از نظر نوع کروموزوم جنسی موجود در هستهشان ناهمگن هستند: 50% از Cها دارای کروموزوم X و 50% دیگر Cs دارای کروموزوم Y هستند. جنسیت کودک توسط "جنس" اسپرم:
اسپرمها (بر خلاف تخممرغ) از نظر نوع کروموزوم جنسی موجود در هستهشان ناهمگن هستند: 50% از Cها دارای کروموزوم X و 50% دیگر Cs دارای کروموزوم Y هستند. جنسیت کودک توسط "جنس" اسپرم:
 لقاح در انسان، نوع لقاح تک اسپرمی: تنها یک اسپرم می تواند وارد تخمک شود. زمان بهینه برای لقاح 24 ساعت اول پس از تخمک گذاری است (اگرچه تخمک II ممکن است برای مدتی توانایی لقاح را حفظ کند). لقاح به طور معمول در آمپول لوله فالوپ رخ می دهد.
لقاح در انسان، نوع لقاح تک اسپرمی: تنها یک اسپرم می تواند وارد تخمک شود. زمان بهینه برای لقاح 24 ساعت اول پس از تخمک گذاری است (اگرچه تخمک II ممکن است برای مدتی توانایی لقاح را حفظ کند). لقاح به طور معمول در آمپول لوله فالوپ رخ می دهد.
 مراحل لقاح 1. برهمکنش دور و همگرایی گامت ها. 2. تعامل تماس و فعال شدن تخم. 3. ورود اسپرم به تخمک و آمیختگی متعاقب آن - سنگامی.
مراحل لقاح 1. برهمکنش دور و همگرایی گامت ها. 2. تعامل تماس و فعال شدن تخم. 3. ورود اسپرم به تخمک و آمیختگی متعاقب آن - سنگامی.
 لقاح 1، 2، 3، 4 - مراحل واکنش آکروزومی 5 - زونا زونا 6 - فضای پریویتلین 7 - غشای پلاسمایی 8 - گرانول قشری 8 الف - واکنش قشری 9 - ورود اسپرم به تخمک 10 - واکنش ناحیه ای
لقاح 1، 2، 3، 4 - مراحل واکنش آکروزومی 5 - زونا زونا 6 - فضای پریویتلین 7 - غشای پلاسمایی 8 - گرانول قشری 8 الف - واکنش قشری 9 - ورود اسپرم به تخمک 10 - واکنش ناحیه ای
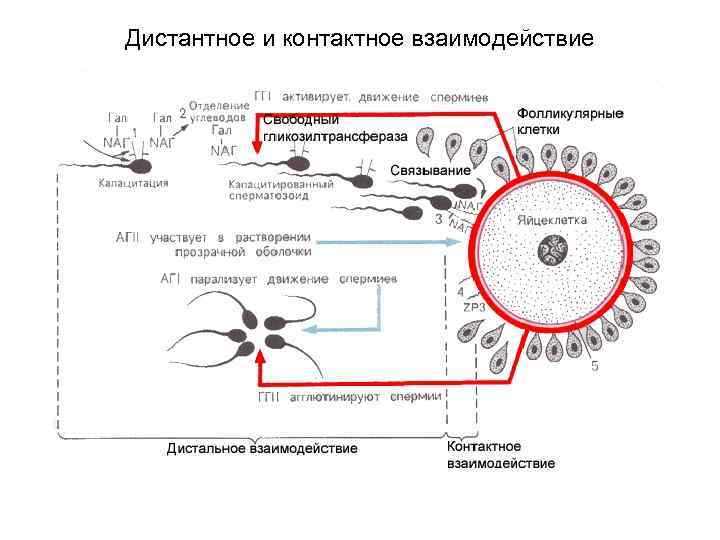
 I. نزدیک شدن و تعامل دور سلول های زایا Oocyte II به آرامی از تخمدان به سمت رحم حرکت می کند. این به طور منفعل اتفاق می افتد - به لطف جریان مخاط. این جریان در اثر کوبیدن مژک های سلول های مژک دار و انقباضات تونیک لوله های فالوپ (تحت عمل پروژسترون) ایجاد می شود. در واژن، تحرک ذاتی اسپرم (Cz) به دلیل محیط اسیدی موجود در اینجا کم است. آنها عمدتاً منفعلانه وارد رحم می شوند - به دلیل انقباضات تونیک دستگاه تناسلی زن. سپس بخشی از C3 نیز، عمدتاً غیرفعال، به لوله های فالوپ می رسد. ظرفیت، یعنی فعال شدن متابولیسم و تحرک C3: C3 به شدت افزایش می یابد و غشاهای C3 در ناحیه سر، گلیکوپروتئین های سطحی را از دست می دهند و بنابراین توانایی اتصال به زونا شفاف را به دست می آورند. ظرفیت سازی توسط gynogamones II ترشح شده توسط تخمک آغاز می شود. حرکت فعال C3 (رئوتاکسی، کموتاکسی) حرکت C3 عمدتاً فعال می شود و با کوبیدن تاژک های آنها ایجاد می شود. در این حالت، اسپرم ها به طور همزمان به جلو حرکت می کنند و حول محور خود می چرخند.
I. نزدیک شدن و تعامل دور سلول های زایا Oocyte II به آرامی از تخمدان به سمت رحم حرکت می کند. این به طور منفعل اتفاق می افتد - به لطف جریان مخاط. این جریان در اثر کوبیدن مژک های سلول های مژک دار و انقباضات تونیک لوله های فالوپ (تحت عمل پروژسترون) ایجاد می شود. در واژن، تحرک ذاتی اسپرم (Cz) به دلیل محیط اسیدی موجود در اینجا کم است. آنها عمدتاً منفعلانه وارد رحم می شوند - به دلیل انقباضات تونیک دستگاه تناسلی زن. سپس بخشی از C3 نیز، عمدتاً غیرفعال، به لوله های فالوپ می رسد. ظرفیت، یعنی فعال شدن متابولیسم و تحرک C3: C3 به شدت افزایش می یابد و غشاهای C3 در ناحیه سر، گلیکوپروتئین های سطحی را از دست می دهند و بنابراین توانایی اتصال به زونا شفاف را به دست می آورند. ظرفیت سازی توسط gynogamones II ترشح شده توسط تخمک آغاز می شود. حرکت فعال C3 (رئوتاکسی، کموتاکسی) حرکت C3 عمدتاً فعال می شود و با کوبیدن تاژک های آنها ایجاد می شود. در این حالت، اسپرم ها به طور همزمان به جلو حرکت می کنند و حول محور خود می چرخند.
 II. برهمکنش تماسی سلولهای زاینده اتصال سلولها با رسیدن به تخمک II، Cهای متعددی به غشاهای آن متصل میشوند (به دلیل تعامل گیرندههای خاص). در همان زمان، به دلیل کوبیدن تاژک C3، تخمک شروع به چرخش حول محور خود می کند. واکنش آکروزومی در Cs متصل، یک واکنش آکروزومی ایجاد میشود: بخشهای قدامی پلاسمولما و غشای آکروزوم پاره میشوند که باعث آزاد شدن آنزیمهای آکروزومی میشود: هیالورونیداز سلولهای غشای دانهای را جدا میکند و آنزیم تریپسین مانند آکروزین و تعدادی دیگر. آنزیم ها زونا پلوسیدا را در محل عبور Cs حل می کنند.
II. برهمکنش تماسی سلولهای زاینده اتصال سلولها با رسیدن به تخمک II، Cهای متعددی به غشاهای آن متصل میشوند (به دلیل تعامل گیرندههای خاص). در همان زمان، به دلیل کوبیدن تاژک C3، تخمک شروع به چرخش حول محور خود می کند. واکنش آکروزومی در Cs متصل، یک واکنش آکروزومی ایجاد میشود: بخشهای قدامی پلاسمولما و غشای آکروزوم پاره میشوند که باعث آزاد شدن آنزیمهای آکروزومی میشود: هیالورونیداز سلولهای غشای دانهای را جدا میکند و آنزیم تریپسین مانند آکروزین و تعدادی دیگر. آنزیم ها زونا پلوسیدا را در محل عبور Cs حل می کنند.
 III. نفوذ اسپرم به داخل تخمک هسته Cs و سانتریول ها به داخل تخمک نفوذ می کنند. پس از آن، یک واکنش قشری در تخمک II در عرض چند ثانیه ایجاد می شود - محتوای گرانول های قشر به خارج از سلول خارج می شود و یک غشای لقاح تشکیل می شود. علاوه بر این، gynogamones I توسط تخمک ترشح می شود که باعث آگلوتیناسیون اسپرم های باقی مانده می شود. تکمیل میوز همزمان مراحل تقسیم دوم میوز (متافاز، آنافاز، تلوفاز) می گذرد. این باعث افزایش تعداد اجسام کاهشی یا قطبی در زیر زونا شفاف می شود.
III. نفوذ اسپرم به داخل تخمک هسته Cs و سانتریول ها به داخل تخمک نفوذ می کنند. پس از آن، یک واکنش قشری در تخمک II در عرض چند ثانیه ایجاد می شود - محتوای گرانول های قشر به خارج از سلول خارج می شود و یک غشای لقاح تشکیل می شود. علاوه بر این، gynogamones I توسط تخمک ترشح می شود که باعث آگلوتیناسیون اسپرم های باقی مانده می شود. تکمیل میوز همزمان مراحل تقسیم دوم میوز (متافاز، آنافاز، تلوفاز) می گذرد. این باعث افزایش تعداد اجسام کاهشی یا قطبی در زیر زونا شفاف می شود.
 آماده سازی زیگوت برای تکه تکه شدن همگرایی هسته ها در زیگوت حاصل، هسته Cs متورم می شود (به یک پیش هسته نر تبدیل می شود و به پیش هسته ماده نزدیک می شود (هسته های به هم پیوسته را سینکاریون می نامند) اما با آن ادغام نمی شود. دو برابر شدن DNA و سانتریول ها مولکول های DNA (در پیش هسته ها) و مولکول هایی که از Cs آمده اند، سانتریول های مضاعف هستند این فرآیندها برای حدود یک روز ادامه می یابد.
آماده سازی زیگوت برای تکه تکه شدن همگرایی هسته ها در زیگوت حاصل، هسته Cs متورم می شود (به یک پیش هسته نر تبدیل می شود و به پیش هسته ماده نزدیک می شود (هسته های به هم پیوسته را سینکاریون می نامند) اما با آن ادغام نمی شود. دو برابر شدن DNA و سانتریول ها مولکول های DNA (در پیش هسته ها) و مولکول هایی که از Cs آمده اند، سانتریول های مضاعف هستند این فرآیندها برای حدود یک روز ادامه می یابد.
 آغاز اولین تقسیم میتوزی تشکیل یک ستاره تک والد اولین تقسیم میتوزی شامل دو پیش هسته است که با هم ادغام نشده اند: پوسته آنها از بین می رود و کروموزوم ها متراکم می شوند و یک ستاره تک والد را در متافاز تشکیل می دهند. در تمام این مدت، پیشرفت آهسته غیرفعال تخمک II، و سپس زیگوت، در امتداد لوله فالوپ تا رحم ادامه دارد.
آغاز اولین تقسیم میتوزی تشکیل یک ستاره تک والد اولین تقسیم میتوزی شامل دو پیش هسته است که با هم ادغام نشده اند: پوسته آنها از بین می رود و کروموزوم ها متراکم می شوند و یک ستاره تک والد را در متافاز تشکیل می دهند. در تمام این مدت، پیشرفت آهسته غیرفعال تخمک II، و سپس زیگوت، در امتداد لوله فالوپ تا رحم ادامه دارد.
 شکل - زیگوت انسانی در مرحله سینکاریون. در شکل، یک غشای لقاح متراکم در اطراف زیگوت قابل مشاهده است (1). ساختار سلولی ندارد، زیرا از زونا پلوسیدا می آید. در داخل زیگوت دو هسته- پیش هسته (2) با حجم تقریبا مساوی وجود دارد. در نتیجه، یک هسته بسیار کوچک و متراکم اسپرم، در نتیجه تراکم زدایی و تورم کروموزوم، به اندازه هسته تخمک می رسد. هر یک از این هسته ها شامل مجموعه ای از کروموزوم هاپلوئید یعنی هر کدام 23 کروموزوم است. از آنجایی که هسته ها قبلاً در تماس هستند (که سیگنالی برای شروع میتوز است)، تکثیر DNA قبلاً در آنها رخ داده است و کروموزوم ها دو کروماتید شده اند.
شکل - زیگوت انسانی در مرحله سینکاریون. در شکل، یک غشای لقاح متراکم در اطراف زیگوت قابل مشاهده است (1). ساختار سلولی ندارد، زیرا از زونا پلوسیدا می آید. در داخل زیگوت دو هسته- پیش هسته (2) با حجم تقریبا مساوی وجود دارد. در نتیجه، یک هسته بسیار کوچک و متراکم اسپرم، در نتیجه تراکم زدایی و تورم کروموزوم، به اندازه هسته تخمک می رسد. هر یک از این هسته ها شامل مجموعه ای از کروموزوم هاپلوئید یعنی هر کدام 23 کروموزوم است. از آنجایی که هسته ها قبلاً در تماس هستند (که سیگنالی برای شروع میتوز است)، تکثیر DNA قبلاً در آنها رخ داده است و کروموزوم ها دو کروماتید شده اند.
 شکاف شکاف زیگوت کامل (هولوبلاستیک)، ناهموار (2 نوع بلاستومر: کوچک، روشن و بزرگتر، تاریک)، ناهمزمان، آهسته است. صفحه اولین خرد کردن از بدنه های کاهش می گذرد.
شکاف شکاف زیگوت کامل (هولوبلاستیک)، ناهموار (2 نوع بلاستومر: کوچک، روشن و بزرگتر، تاریک)، ناهمزمان، آهسته است. صفحه اولین خرد کردن از بدنه های کاهش می گذرد.

 تقسیم کردن. خصوصیات کلی حفظ غشای لقاح در طی 2-4 روز، تکه تکه شدن رخ می دهد، یعنی مجموعه ای از تقسیمات میتوزی بدون دوره های رشد سلول های دختر. رشد سلولی با این واقعیت که یک پوسته لقاح متراکم در اطراف جنین حفظ می شود، مانع از رشد سلولی می شود که از هجوم مواد مغذی از خارج نیز جلوگیری می کند (فعالیت زندگی با هزینه ذخایر تخمک پشتیبانی می شود). و افزایش بسیار در اندازه جنین. کاهش اندازه سلول ها با توجه به موارد فوق، سلول های به طور فزاینده ای کوچکتر تشکیل می شوند و حجم کل جنین افزایش نمی یابد.
تقسیم کردن. خصوصیات کلی حفظ غشای لقاح در طی 2-4 روز، تکه تکه شدن رخ می دهد، یعنی مجموعه ای از تقسیمات میتوزی بدون دوره های رشد سلول های دختر. رشد سلولی با این واقعیت که یک پوسته لقاح متراکم در اطراف جنین حفظ می شود، مانع از رشد سلولی می شود که از هجوم مواد مغذی از خارج نیز جلوگیری می کند (فعالیت زندگی با هزینه ذخایر تخمک پشتیبانی می شود). و افزایش بسیار در اندازه جنین. کاهش اندازه سلول ها با توجه به موارد فوق، سلول های به طور فزاینده ای کوچکتر تشکیل می شوند و حجم کل جنین افزایش نمی یابد.

 تشکیل بلاستوسیست پس از 4، 5 تا 5 روز، یک بلاستوسیست تشکیل می شود - یک وزیکول جنینی پر از مایع. اجزای بلاستوسیست به شرح زیر است: بلاستومرهای سبک کوچکتر تروفوبلاست را تشکیل می دهند. تروفوبلاست - یک دیواره تک لایه ای از سلول های سبک کوچک (متعاقباً یک اندام خارج جنینی، کوریون، از تروفوبلاست ایجاد می شود). تیرههای بزرگتر - جنینوبلاست، یا توده سلولی داخلی - تجمع بلاستومرهای تیره بزرگ به شکل یک گره در سطح داخلی تروفوبلاست در یکی از قطبها. بلاستوکول یک حفره پر از مایع است.
تشکیل بلاستوسیست پس از 4، 5 تا 5 روز، یک بلاستوسیست تشکیل می شود - یک وزیکول جنینی پر از مایع. اجزای بلاستوسیست به شرح زیر است: بلاستومرهای سبک کوچکتر تروفوبلاست را تشکیل می دهند. تروفوبلاست - یک دیواره تک لایه ای از سلول های سبک کوچک (متعاقباً یک اندام خارج جنینی، کوریون، از تروفوبلاست ایجاد می شود). تیرههای بزرگتر - جنینوبلاست، یا توده سلولی داخلی - تجمع بلاستومرهای تیره بزرگ به شکل یک گره در سطح داخلی تروفوبلاست در یکی از قطبها. بلاستوکول یک حفره پر از مایع است.


 رشد بلاستوسیست به دلیل جذب مایع از حفره رحم توسط تروفوبلاست، حجم وزیکول کمی افزایش می یابد. برآمدگی هایی در تروفوبلاست ظاهر می شود که به تدریج غشای لقاح اطراف جنین را از بین می برد. در نتیجه چند ساعت قبل از لانه گزینی، جنین این غشاء را از دست می دهد. پس از آن، پوسته دیگر از افزایش اندازه جنین جلوگیری نمی کند و از آن به بعد، چرخه سلولی میتوزی طبیعی می شود، یعنی شامل فاز رشد می شود. بنابراین، در افزایش بعدی در جرم جنین بسیار سریعتر اتفاق می افتد.
رشد بلاستوسیست به دلیل جذب مایع از حفره رحم توسط تروفوبلاست، حجم وزیکول کمی افزایش می یابد. برآمدگی هایی در تروفوبلاست ظاهر می شود که به تدریج غشای لقاح اطراف جنین را از بین می برد. در نتیجه چند ساعت قبل از لانه گزینی، جنین این غشاء را از دست می دهد. پس از آن، پوسته دیگر از افزایش اندازه جنین جلوگیری نمی کند و از آن به بعد، چرخه سلولی میتوزی طبیعی می شود، یعنی شامل فاز رشد می شود. بنابراین، در افزایش بعدی در جرم جنین بسیار سریعتر اتفاق می افتد.
 بلاستوسیست تا روز 67 e در حفره رحم آزاد می ماند. آر. ، سپس در ترشحات مخاطی غدد رحمی در سطح آندومتر فرو می رود.
بلاستوسیست تا روز 67 e در حفره رحم آزاد می ماند. آر. ، سپس در ترشحات مخاطی غدد رحمی در سطح آندومتر فرو می رود.
 لانه گزینی - فرآیند نفوذ جنین به غشای مخاطی دیواره رحم (آندومتر) و ایجاد پیوندهای نزدیک با عروق خونی آن - از روز 6-7 آغاز می شود. این به دو مرحله تقسیم می شود - چسبندگی و تهاجم تحت تأثیر ترشح رحم، زونا پلوسیدا حل می شود و بلاستوسیست به آندومتر می چسبد (چسبندگی). سلول های تروفوبلاست به سرعت رشد می کنند، آنزیم های لیزوزومی ترشح می کنند و بافت آندومتر را تخریب می کنند. معمولا لانه گزینی در آندومتر دیواره خلفی رحم اتفاق می افتد.
لانه گزینی - فرآیند نفوذ جنین به غشای مخاطی دیواره رحم (آندومتر) و ایجاد پیوندهای نزدیک با عروق خونی آن - از روز 6-7 آغاز می شود. این به دو مرحله تقسیم می شود - چسبندگی و تهاجم تحت تأثیر ترشح رحم، زونا پلوسیدا حل می شود و بلاستوسیست به آندومتر می چسبد (چسبندگی). سلول های تروفوبلاست به سرعت رشد می کنند، آنزیم های لیزوزومی ترشح می کنند و بافت آندومتر را تخریب می کنند. معمولا لانه گزینی در آندومتر دیواره خلفی رحم اتفاق می افتد.

 تروفوبلاست با شروع لانه گزینی (به محض تماس تروفوبلاست با اپیتلیوم آندومتر)، سلول های تروفوبلاست شروع به تقسیم فعال می کنند، سلول های دختر ادغام می شوند و یک توده سیتوپلاسمی منفرد را تشکیل می دهند که حاوی هسته های زیادی و بدون مرز سلولی است و یک سمپلاستو-( syncytio)-trophoblast (برگ بیرونی) - یک سمپلاست چند هسته ای که در نتیجه ادغام تعداد زیادی سلول ایجاد می شود. و سیتوتروفوبلاست - (برگ داخلی) (ساختار سلولی را حفظ می کند).
تروفوبلاست با شروع لانه گزینی (به محض تماس تروفوبلاست با اپیتلیوم آندومتر)، سلول های تروفوبلاست شروع به تقسیم فعال می کنند، سلول های دختر ادغام می شوند و یک توده سیتوپلاسمی منفرد را تشکیل می دهند که حاوی هسته های زیادی و بدون مرز سلولی است و یک سمپلاستو-( syncytio)-trophoblast (برگ بیرونی) - یک سمپلاست چند هسته ای که در نتیجه ادغام تعداد زیادی سلول ایجاد می شود. و سیتوتروفوبلاست - (برگ داخلی) (ساختار سلولی را حفظ می کند).

 پس از کاشت، سمپلاستوتروفوبلاست محصولات پوسیدگی بافت را جذب می کند - یک نوع تغذیه هیستوتروف (دو هفته اول). بنابراین، نقش سمپلاستوتروفوبلاست این است که: به دلیل عملکرد آنزیم های لیزوزومی، تهاجم جنین به دیواره رحم را تضمین می کند. تغذیه لازم برای رشد جنین در دو هفته اول زندگی را فراهم می کند و محصولات پوسیدگی بافت های رحم را جذب می کند (نوع تغذیه هیستوتروف). عملکرد غدد درون ریز را انجام می دهد (stb - محل تشکیل هورمون ها از جمله hCG (هورمونی که تشکیل پروژسترون توسط جسم زرد را تحریک می کند - در خون یا ادرار زنان از روز دهم بارداری تعیین می شود و بر اساس آزمایش بارداری).
پس از کاشت، سمپلاستوتروفوبلاست محصولات پوسیدگی بافت را جذب می کند - یک نوع تغذیه هیستوتروف (دو هفته اول). بنابراین، نقش سمپلاستوتروفوبلاست این است که: به دلیل عملکرد آنزیم های لیزوزومی، تهاجم جنین به دیواره رحم را تضمین می کند. تغذیه لازم برای رشد جنین در دو هفته اول زندگی را فراهم می کند و محصولات پوسیدگی بافت های رحم را جذب می کند (نوع تغذیه هیستوتروف). عملکرد غدد درون ریز را انجام می دهد (stb - محل تشکیل هورمون ها از جمله hCG (هورمونی که تشکیل پروژسترون توسط جسم زرد را تحریک می کند - در خون یا ادرار زنان از روز دهم بارداری تعیین می شود و بر اساس آزمایش بارداری).

 پس از غوطه ور شدن کامل جنین، نقص در آندومتر ابتدا با خون منعقد شده پر می شود، سپس با بافت همبند بیش از حد رشد می کند و تا 12-13 روز با اپیتلیوم پوشانده می شود. شکاف هایی در اطراف جنین ایجاد می شود که با خون مادر پر شده است. در سلول های استرومای آندومتر، به اصطلاح. واکنش decidual: عروق رشد می کنند، تورم ظاهر می شود، حجم سلول ها افزایش می یابد و گلیکوژن و لیپیدها جمع می شوند.
پس از غوطه ور شدن کامل جنین، نقص در آندومتر ابتدا با خون منعقد شده پر می شود، سپس با بافت همبند بیش از حد رشد می کند و تا 12-13 روز با اپیتلیوم پوشانده می شود. شکاف هایی در اطراف جنین ایجاد می شود که با خون مادر پر شده است. در سلول های استرومای آندومتر، به اصطلاح. واکنش decidual: عروق رشد می کنند، تورم ظاهر می شود، حجم سلول ها افزایش می یابد و گلیکوژن و لیپیدها جمع می شوند.
 گاسترولاسیون (هفته 2) گاسترولاسیون در انسان در دو مرحله رخ می دهد. 1) شکافتن (لایه برداری) در روز هفتم، - همزمان با کاشت، یک اپی بلاست و یک هیپوبلاست تشکیل می شود. 2) مهاجرت در روز 14 - 15، یک رگه اولیه، یک ندول سر، تشکیل می شود. 3) بین این دو مرحله، تشکیل اندام های خارج جنینی لازم برای رشد موفقیت آمیز جنین وجود دارد.
گاسترولاسیون (هفته 2) گاسترولاسیون در انسان در دو مرحله رخ می دهد. 1) شکافتن (لایه برداری) در روز هفتم، - همزمان با کاشت، یک اپی بلاست و یک هیپوبلاست تشکیل می شود. 2) مهاجرت در روز 14 - 15، یک رگه اولیه، یک ندول سر، تشکیل می شود. 3) بین این دو مرحله، تشکیل اندام های خارج جنینی لازم برای رشد موفقیت آمیز جنین وجود دارد.
 گاسترولاسیون فاز 1 - لایه لایه شدن - در روز هفتم جنین زایی. جنین به اپی بلاست و هیپوبلاست تقسیم می شود - آندودرم خارج جنینی (زرده) آینده.
گاسترولاسیون فاز 1 - لایه لایه شدن - در روز هفتم جنین زایی. جنین به اپی بلاست و هیپوبلاست تقسیم می شود - آندودرم خارج جنینی (زرده) آینده.

 سلول های مزودرم از اپی بلاست خارج می شوند و باعث ایجاد مزودرم خارج جنینی می شوند. مزودرم خارج جنینی یک ساختار اسفنجی شکل می دهد که توسط رشته های سلولی تشکیل شده است. به تدریج دیواره های آمنیوتیک و وزیکول زرده را بیش از حد رشد می کند و غشای آمنیوتیک (از داخل با اکتودرم خارج جنینی، از بیرون - توسط مزودرم خارج جنینی) و کیسه زرده (از داخل با پوشش خارج جنینی پوشانده شده است) تشکیل می دهد. اندودرم، از خارج - توسط مزودرم خارج جنینی). هنگامی که مزودرم به سمت تروفوبلاست رشد می کند، یک غشای کوریونی تشکیل می شود (مزودرم خارج جنینی + سیتو- و سمپلاستوتروفوبلاست) و حفره های کوچک در یک حفره منفرد کوریون (مترادف - کل خارج جنینی) ادغام می شوند.
سلول های مزودرم از اپی بلاست خارج می شوند و باعث ایجاد مزودرم خارج جنینی می شوند. مزودرم خارج جنینی یک ساختار اسفنجی شکل می دهد که توسط رشته های سلولی تشکیل شده است. به تدریج دیواره های آمنیوتیک و وزیکول زرده را بیش از حد رشد می کند و غشای آمنیوتیک (از داخل با اکتودرم خارج جنینی، از بیرون - توسط مزودرم خارج جنینی) و کیسه زرده (از داخل با پوشش خارج جنینی پوشانده شده است) تشکیل می دهد. اندودرم، از خارج - توسط مزودرم خارج جنینی). هنگامی که مزودرم به سمت تروفوبلاست رشد می کند، یک غشای کوریونی تشکیل می شود (مزودرم خارج جنینی + سیتو- و سمپلاستوتروفوبلاست) و حفره های کوچک در یک حفره منفرد کوریون (مترادف - کل خارج جنینی) ادغام می شوند.


 تشکیل کیسه زرده رشد سلول های هیپوبلاست مزودرم خارج جنینی مهاجرت کرده و ادغام می شوند و دیواره کیسه زرده را تشکیل می دهند. Epiblast - پایین کیسه آمنیوتیک؛ هیپوبلاست سقف کیسه زرده است. مزودرم خارج جنینی (مزانشیم اولیه) یک ساختار اسفنجی را تشکیل می دهد.
تشکیل کیسه زرده رشد سلول های هیپوبلاست مزودرم خارج جنینی مهاجرت کرده و ادغام می شوند و دیواره کیسه زرده را تشکیل می دهند. Epiblast - پایین کیسه آمنیوتیک؛ هیپوبلاست سقف کیسه زرده است. مزودرم خارج جنینی (مزانشیم اولیه) یک ساختار اسفنجی را تشکیل می دهد.
 جنین از طریق ساقه آمنیوتیک تشکیل شده توسط مزودرم خارج جنینی به کوریون در حال رشد متصل می شود. مزودرم خارج جنینی به بیرون از تروفوبلاست رشد می کند و پرزهای کوریونی اولیه ثانویه می شوند (در روز 12-13).
جنین از طریق ساقه آمنیوتیک تشکیل شده توسط مزودرم خارج جنینی به کوریون در حال رشد متصل می شود. مزودرم خارج جنینی به بیرون از تروفوبلاست رشد می کند و پرزهای کوریونی اولیه ثانویه می شوند (در روز 12-13).
 1 - اپی بلاست ژرمینال. 2- اکتودرم آمنیوتیک. 3- حفره آمنیوتیک (بین دو ورقه قبلی). 4 - وزیکول زرده (از هیپوبلاست تشکیل شده است). سطوح رشد بیش از حد با مزودرم خارج جنینی: 5 A - سطح خارجی وزیکول آمنیوتیک. 5 B - سطح بیرونی کیسه زرده؛ 5 V - سطح داخلی تروفوبلاست. 6- کوریون: این تروفوبلاست همراه با مزودرم است که به آن رشد کرده است. 7- پای آمنیوتیک: از مزودرم خارج جنینی تشکیل شده و جنین را به کوریون متصل می کند.
1 - اپی بلاست ژرمینال. 2- اکتودرم آمنیوتیک. 3- حفره آمنیوتیک (بین دو ورقه قبلی). 4 - وزیکول زرده (از هیپوبلاست تشکیل شده است). سطوح رشد بیش از حد با مزودرم خارج جنینی: 5 A - سطح خارجی وزیکول آمنیوتیک. 5 B - سطح بیرونی کیسه زرده؛ 5 V - سطح داخلی تروفوبلاست. 6- کوریون: این تروفوبلاست همراه با مزودرم است که به آن رشد کرده است. 7- پای آمنیوتیک: از مزودرم خارج جنینی تشکیل شده و جنین را به کوریون متصل می کند.

 هفته دوم جنین زایی دوره "دو" نامیده می شود: دو لایه تشکیل می شود - اپی بلاست و هیپوبلاست که دیسک ژرمینال را تشکیل می دهند. دو وزیکول ایجاد می شود - آمنیوتیک و ویتلین. دو لایه تروفوبلاست متمایز می شود - سیتو- و سمپلاستوتروفوبلاست. در پایان این هفته، تروفوبلاست برآمدگی های شاخه ای را تشکیل می دهد - پرزهای اولیه، که پس از آن قبلاً کوریون (غشای پرز) نامیده می شود.
هفته دوم جنین زایی دوره "دو" نامیده می شود: دو لایه تشکیل می شود - اپی بلاست و هیپوبلاست که دیسک ژرمینال را تشکیل می دهند. دو وزیکول ایجاد می شود - آمنیوتیک و ویتلین. دو لایه تروفوبلاست متمایز می شود - سیتو- و سمپلاستوتروفوبلاست. در پایان این هفته، تروفوبلاست برآمدگی های شاخه ای را تشکیل می دهد - پرزهای اولیه، که پس از آن قبلاً کوریون (غشای پرز) نامیده می شود.

 بنابراین، در پایان هفته اول - آغاز هفته دوم رشد دوران بارداری، فرآیندهای بسیار مهم و همزمان رخ می دهد: لانه گزینی. مرحله اول گاسترولاسیون؛ ظهور مزودرم خارج جنینی و رشد اندام های خارج جنینی: آمنیون، کیسه زرده، کوریون. در این راستا، پایان هفته اول زندگی جنین به دوره بحرانی (تعیین کننده) رشد (7-8 روز قبل از آن) اشاره دارد.
بنابراین، در پایان هفته اول - آغاز هفته دوم رشد دوران بارداری، فرآیندهای بسیار مهم و همزمان رخ می دهد: لانه گزینی. مرحله اول گاسترولاسیون؛ ظهور مزودرم خارج جنینی و رشد اندام های خارج جنینی: آمنیون، کیسه زرده، کوریون. در این راستا، پایان هفته اول زندگی جنین به دوره بحرانی (تعیین کننده) رشد (7-8 روز قبل از آن) اشاره دارد.
 هفته سوم رشد مرحله دوم گاسترولاسیون - مهاجرت در روز 14-15 جنین زایی اتفاق می افتد.
هفته سوم رشد مرحله دوم گاسترولاسیون - مهاجرت در روز 14-15 جنین زایی اتفاق می افتد.

 سلولهایی که در ناحیه گره هنسن مهاجرت میکنند، یک فرآیند کوردومزودرم را تشکیل میدهند که در زیر اپی بلاست به سمت انتهای سر در امتداد خط وسط جنین رشد میکند. سلول های این فرآیند به مزودرم سر و نوتوکورد تمایز می یابند. نقش نوتوکورد در روند رشد: ✴نوتوکورد محور ستون فقرات آینده است. و صفحه عصبی و لوله عصبی تشکیل می شود. ✴نوکلئوس پالپوزوس
سلولهایی که در ناحیه گره هنسن مهاجرت میکنند، یک فرآیند کوردومزودرم را تشکیل میدهند که در زیر اپی بلاست به سمت انتهای سر در امتداد خط وسط جنین رشد میکند. سلول های این فرآیند به مزودرم سر و نوتوکورد تمایز می یابند. نقش نوتوکورد در روند رشد: ✴نوتوکورد محور ستون فقرات آینده است. و صفحه عصبی و لوله عصبی تشکیل می شود. ✴نوکلئوس پالپوزوس


 مزانشیم سلولهای مزانشیم از تمام لایههای زایایی خارج میشوند، بهطور فعال از لایه احشایی اسپلانکنوتوم. چندین نوع بافت و ساختار از مزانشیم تشکیل می شود: خون. انواع بافت همبند؛ بافت ماهیچه صاف، عروق خونی و اندوکارد قلب؛ میکروگلیای سیستم عصبی مرکزی
مزانشیم سلولهای مزانشیم از تمام لایههای زایایی خارج میشوند، بهطور فعال از لایه احشایی اسپلانکنوتوم. چندین نوع بافت و ساختار از مزانشیم تشکیل می شود: خون. انواع بافت همبند؛ بافت ماهیچه صاف، عروق خونی و اندوکارد قلب؛ میکروگلیای سیستم عصبی مرکزی


 پرزهای کوریونیک سوم رگ های خونی جنین به پرزهای کوریونی تبدیل می شوند که به حالت سوم تبدیل می شوند (در هفته سوم رشد ظاهر می شوند).
پرزهای کوریونیک سوم رگ های خونی جنین به پرزهای کوریونی تبدیل می شوند که به حالت سوم تبدیل می شوند (در هفته سوم رشد ظاهر می شوند).

 چین تنه از روز بیستم - آغاز جدا شدن بدن جنین از غشاهای خارج جنینی - چین تنه تشکیل می شود. در نتیجه تشکیل چین های تنه، بدن جنین از اندام های موقتی بالا می رود و از آنها جدا می شود. در این مورد، جنین، همانطور که بود، به یک لوله می پیچد. در عین حال، این منجر به تشکیل یک لوله روده از آندودرم روده می شود که از آندودرم خارج جنینی کیسه زرده جدا می شود.
چین تنه از روز بیستم - آغاز جدا شدن بدن جنین از غشاهای خارج جنینی - چین تنه تشکیل می شود. در نتیجه تشکیل چین های تنه، بدن جنین از اندام های موقتی بالا می رود و از آنها جدا می شود. در این مورد، جنین، همانطور که بود، به یک لوله می پیچد. در عین حال، این منجر به تشکیل یک لوله روده از آندودرم روده می شود که از آندودرم خارج جنینی کیسه زرده جدا می شود.

 آمنیون یک حفره بسته در اطراف جنین ایجاد می کند. توابع: ایجاد یک محیط آبی با ترکیب شیمیایی خاص و فشار برای توسعه آزاد، محافظت در برابر تنش های مکانیکی و گرانشی. به تدریج، حفره رشد می کند. در هفته هفتم رشد، مزودرم آمنیوتیک با مزودرم کوریونی (غشای آمنیو کوریونی) تماس پیدا می کند. علاوه بر این، اپیتلیوم آمنیوتیک ساقه آمنیوتیک را بیش از حد رشد می کند.
آمنیون یک حفره بسته در اطراف جنین ایجاد می کند. توابع: ایجاد یک محیط آبی با ترکیب شیمیایی خاص و فشار برای توسعه آزاد، محافظت در برابر تنش های مکانیکی و گرانشی. به تدریج، حفره رشد می کند. در هفته هفتم رشد، مزودرم آمنیوتیک با مزودرم کوریونی (غشای آمنیو کوریونی) تماس پیدا می کند. علاوه بر این، اپیتلیوم آمنیوتیک ساقه آمنیوتیک را بیش از حد رشد می کند.

 آمنیون تا لحظه تولد (مثانه جنین) عمل می کند. تا پایان بارداری، 1-1.5 لیتر مایع آمنیوتیک (مایع آمنیوتیک) پر می شود.
آمنیون تا لحظه تولد (مثانه جنین) عمل می کند. تا پایان بارداری، 1-1.5 لیتر مایع آمنیوتیک (مایع آمنیوتیک) پر می شود.
 کیسه زرده فقط در مراحل اولیه رشد عمل می کند: محلی سازی اولین کانون های خون سازی و تشکیل رگ های خونی و سلول های زایای اولیه. پس از 7-8 هفته، دچار پسرفت می شود، به شکل طناب سلولی در بند ناف باقی می ماند و رگ های خونی را به جفت هدایت می کند. بر خلاف پرندگان، در انسان (و سایر پستانداران) کیسه زرده عملاً حاوی زرده نیست، اما با مایع سروزی پر شده است.
کیسه زرده فقط در مراحل اولیه رشد عمل می کند: محلی سازی اولین کانون های خون سازی و تشکیل رگ های خونی و سلول های زایای اولیه. پس از 7-8 هفته، دچار پسرفت می شود، به شکل طناب سلولی در بند ناف باقی می ماند و رگ های خونی را به جفت هدایت می کند. بر خلاف پرندگان، در انسان (و سایر پستانداران) کیسه زرده عملاً حاوی زرده نیست، اما با مایع سروزی پر شده است.
 آلانتویس در روز 17-18 به شکل یک رشد کوچک کور از دیواره شکمی روده خلفی ایجاد می شود. آلانتوا در ساقه آمنیوتیک رشد می کند، رگ های خونی نافی در دیواره آن ایجاد می شود که آن را به کوریون (رسانای رگ) می رساند. در ماه دوم ه. آر. کاهش می یابد و همراه با بقایای کیسه زرده، یک بند سلولی را به عنوان بخشی از بند ناف تشکیل می دهد.
آلانتویس در روز 17-18 به شکل یک رشد کوچک کور از دیواره شکمی روده خلفی ایجاد می شود. آلانتوا در ساقه آمنیوتیک رشد می کند، رگ های خونی نافی در دیواره آن ایجاد می شود که آن را به کوریون (رسانای رگ) می رساند. در ماه دوم ه. آر. کاهش می یابد و همراه با بقایای کیسه زرده، یک بند سلولی را به عنوان بخشی از بند ناف تشکیل می دهد.
 کوریون - قسمت جنینی جفت در تشکیل کوریون، سه دوره متمایز می شود: پیش پرز (7-8 روز)، دوره تشکیل پرز (تا 50 روز)، دوره لپه (از 50 تا 90 روز). . این توسط صفحه کوریونی (مزودرم خارج جنینی) و برآمدگی های صفحه - پرزهای سوم منشعب پوشیده شده با تروفوبلاست تشکیل می شود.
کوریون - قسمت جنینی جفت در تشکیل کوریون، سه دوره متمایز می شود: پیش پرز (7-8 روز)، دوره تشکیل پرز (تا 50 روز)، دوره لپه (از 50 تا 90 روز). . این توسط صفحه کوریونی (مزودرم خارج جنینی) و برآمدگی های صفحه - پرزهای سوم منشعب پوشیده شده با تروفوبلاست تشکیل می شود.
 تشکیل پرزهای اولیه (پایان هفته دوم) تا اواسط هفته دوم رشد، جنین قبلاً به طور کامل در آندومتر غوطه ور شده است و تروفوبلاست از دو لایه - بیرونی (سمپلاستوتروفوبلاست) و داخلی (سیتوتروفوبلاست) تشکیل شده است. . در پایان این هفته، تروفوبلاست برآمدگی های شاخه ای را تشکیل می دهد - پرزهای اولیه، که پس از آن قبلاً کوریون (غشای پرز) نامیده می شود.
تشکیل پرزهای اولیه (پایان هفته دوم) تا اواسط هفته دوم رشد، جنین قبلاً به طور کامل در آندومتر غوطه ور شده است و تروفوبلاست از دو لایه - بیرونی (سمپلاستوتروفوبلاست) و داخلی (سیتوتروفوبلاست) تشکیل شده است. . در پایان این هفته، تروفوبلاست برآمدگی های شاخه ای را تشکیل می دهد - پرزهای اولیه، که پس از آن قبلاً کوریون (غشای پرز) نامیده می شود.
 تشکیل پرزهای ثانویه (ابتدای هفته سوم) در ابتدای هفته سوم، مزانشیم خارج جنینی به داخل پرزهای کوریون (که همه سطوح از داخل از جمله سطح داخلی تروفوبلاستاکوریون را بیش از حد رشد می کند) نفوذ می کند. بنابراین، استرومای بافت همبند پرزها تشکیل می شود و خود پرزها ثانویه می شوند.
تشکیل پرزهای ثانویه (ابتدای هفته سوم) در ابتدای هفته سوم، مزانشیم خارج جنینی به داخل پرزهای کوریون (که همه سطوح از داخل از جمله سطح داخلی تروفوبلاستاکوریون را بیش از حد رشد می کند) نفوذ می کند. بنابراین، استرومای بافت همبند پرزها تشکیل می شود و خود پرزها ثانویه می شوند.
 تشکیل پرزهای سوم (پایان هفته سوم) تا پایان هفته سوم، عروق خونی اولیه در داخل پرزها از سلول های مزانشیمی تشکیل می شوند که به صورت هموکاپیلاری ساخته شده و با رگ های صفحه مزودرم کوریون در تماس هستند. . پرزهایی که رگ های خونی توسعه یافته دارند، ثالثی نامیده می شوند.
تشکیل پرزهای سوم (پایان هفته سوم) تا پایان هفته سوم، عروق خونی اولیه در داخل پرزها از سلول های مزانشیمی تشکیل می شوند که به صورت هموکاپیلاری ساخته شده و با رگ های صفحه مزودرم کوریون در تماس هستند. . پرزهایی که رگ های خونی توسعه یافته دارند، ثالثی نامیده می شوند.
 بخشی از کوریون که دیواره رحم را از بین می برد و در تشکیل جفت شرکت می کند پرزهای منشعب پیچیده ای را تشکیل می دهد و کوریون پرز نامیده می شود. بقیه سطح کوریون صاف است. بزرگترین پرزهایی که از صفحه کوریونی خارج می شوند پرزهای ساقه نامیده می شوند. پرزهای ساقه به فراوانی منشعب می شوند، کوچکترین شاخه ها پرزهای انتهایی نامیده می شوند. پرزهایی که به لایه بازال آندومتر نفوذ می کنند، پرزهای لنگر نامیده می شوند. معمولا پرزهای ساقه لنگر هستند.
بخشی از کوریون که دیواره رحم را از بین می برد و در تشکیل جفت شرکت می کند پرزهای منشعب پیچیده ای را تشکیل می دهد و کوریون پرز نامیده می شود. بقیه سطح کوریون صاف است. بزرگترین پرزهایی که از صفحه کوریونی خارج می شوند پرزهای ساقه نامیده می شوند. پرزهای ساقه به فراوانی منشعب می شوند، کوچکترین شاخه ها پرزهای انتهایی نامیده می شوند. پرزهایی که به لایه بازال آندومتر نفوذ می کنند، پرزهای لنگر نامیده می شوند. معمولا پرزهای ساقه لنگر هستند.

 جفت انسان جفت تنها اندامی است که از سلول های دو موجود ژنتیکی متفاوت تشکیل شده است. شامل قسمت جنینی (کوریون با پرزها) قسمت مادری - آندومتر تغییر یافته است.
جفت انسان جفت تنها اندامی است که از سلول های دو موجود ژنتیکی متفاوت تشکیل شده است. شامل قسمت جنینی (کوریون با پرزها) قسمت مادری - آندومتر تغییر یافته است.
 جفت شدن دوره ای از جنین زایی است که در طی آن رشد جفت رخ می دهد، یکی از دوره های بحرانی جنین زایی، مربوط به هفته های 3-8 بارداری است. لایه سطحی آندومتر غشای دسیدوال (افتاده) نامیده می شود. بسته به محل نسبت به محل لانه گزینی، موارد زیر وجود دارد: Decidua parietalis (پاریتال) - آندومتر پوشاننده حفره رحم به استثنای محل لانه گزینی. Decidua capsularis (کیسه) - بخشی از آندومتر که جنین در حال رشد را احاطه می کند، یک کپسول در بالای آن تشکیل می دهد و جنین را از حفره رحم جدا می کند (تا هفته شانزدهم) Decidua basalis (اصلی)، قسمت مادری رحم جفت، بخشی از آندومتر که بین جنین و لایه پایه آندومتر قرار دارد. بنابراین، قسمت مادری جفت توسط: صفحه پایه (decidua basalis) آندومتر نشان داده می شود. شکاف پر از خون مادر
جفت شدن دوره ای از جنین زایی است که در طی آن رشد جفت رخ می دهد، یکی از دوره های بحرانی جنین زایی، مربوط به هفته های 3-8 بارداری است. لایه سطحی آندومتر غشای دسیدوال (افتاده) نامیده می شود. بسته به محل نسبت به محل لانه گزینی، موارد زیر وجود دارد: Decidua parietalis (پاریتال) - آندومتر پوشاننده حفره رحم به استثنای محل لانه گزینی. Decidua capsularis (کیسه) - بخشی از آندومتر که جنین در حال رشد را احاطه می کند، یک کپسول در بالای آن تشکیل می دهد و جنین را از حفره رحم جدا می کند (تا هفته شانزدهم) Decidua basalis (اصلی)، قسمت مادری رحم جفت، بخشی از آندومتر که بین جنین و لایه پایه آندومتر قرار دارد. بنابراین، قسمت مادری جفت توسط: صفحه پایه (decidua basalis) آندومتر نشان داده می شود. شکاف پر از خون مادر
 لپه ها پرزهای رو به دسیدوای بازالیس به طور ناهموار توزیع شده اند، در گروه ها - لپه ها. لپه یک واحد ساختاری و عملکردی جفت تشکیل شده است. توسط پرز ساقه و شاخه های آن تشکیل می شود. لپه ها تا حدی توسط سپتوم های بافت همبند (سپتوم جفت) از هم جدا می شوند.
لپه ها پرزهای رو به دسیدوای بازالیس به طور ناهموار توزیع شده اند، در گروه ها - لپه ها. لپه یک واحد ساختاری و عملکردی جفت تشکیل شده است. توسط پرز ساقه و شاخه های آن تشکیل می شود. لپه ها تا حدی توسط سپتوم های بافت همبند (سپتوم جفت) از هم جدا می شوند.

 سد خونی جفتی خون مادر و کودک هرگز با هم مخلوط نمی شوند اجزای سد هماتو جفتی جداکننده خون مادر و جنین: اندوتلیوم مویرگی جنین. غشای پایه در دیواره مویرگ های جنین؛ بافت همبند پرز (با سلول های ماکروفاژ)؛ غشای پایه تروفوبلاست؛ سیتوتروفوبلاست؛ سمپلاستوتروفوبلاست ارتباط بین گردش خون جنین و مادر از طریق بند ناف انجام می شود
سد خونی جفتی خون مادر و کودک هرگز با هم مخلوط نمی شوند اجزای سد هماتو جفتی جداکننده خون مادر و جنین: اندوتلیوم مویرگی جنین. غشای پایه در دیواره مویرگ های جنین؛ بافت همبند پرز (با سلول های ماکروفاژ)؛ غشای پایه تروفوبلاست؛ سیتوتروفوبلاست؛ سمپلاستوتروفوبلاست ارتباط بین گردش خون جنین و مادر از طریق بند ناف انجام می شود
 سد هماتو جفت خون مادر (1) و کودک (8) با هم مخلوط نمی شوند: - اندوتلیوم مویرگی جنین (7). - غشای پایه در دیواره مویرگ های جنین (6)؛ - بافت همبند پرز (5)؛ - غشای پایه تروفوبلاست (4)؛ - سیتوتوروفوبلاست (3)؛ - سمپلاستوتروفوبلاست (2)
سد هماتو جفت خون مادر (1) و کودک (8) با هم مخلوط نمی شوند: - اندوتلیوم مویرگی جنین (7). - غشای پایه در دیواره مویرگ های جنین (6)؛ - بافت همبند پرز (5)؛ - غشای پایه تروفوبلاست (4)؛ - سیتوتوروفوبلاست (3)؛ - سمپلاستوتروفوبلاست (2)

 نفوذپذیری خون جفتی به تعدادی از مواد و پاتوژن ها پاتوژن ها ویروس ها (HIV، سیتومگالوویروس، سرخجه، آبله، آبله مرغان، سرخک، فلج اطفال) باکتری ها (سل، ترپونما) تک یاخته ها (توکسوپلاسما) مواد مخدر و فرآورده های دارویی، نیکوتین، کوکائین، کوکائین، داروهای ضد سرطان دیفنین تتراسایکلین، داروهای بیهوشی، آرام بخش، ضد درد آنتی بادی های فاکتور Rh
نفوذپذیری خون جفتی به تعدادی از مواد و پاتوژن ها پاتوژن ها ویروس ها (HIV، سیتومگالوویروس، سرخجه، آبله، آبله مرغان، سرخک، فلج اطفال) باکتری ها (سل، ترپونما) تک یاخته ها (توکسوپلاسما) مواد مخدر و فرآورده های دارویی، نیکوتین، کوکائین، کوکائین، داروهای ضد سرطان دیفنین تتراسایکلین، داروهای بیهوشی، آرام بخش، ضد درد آنتی بادی های فاکتور Rh
 عملکرد جفت تغذیه ای است (انواع مواد مغذی، الکترولیت ها، ویتامین ها و برخی هورمون ها از بدن مادر به جنین می رسد؛ تنفسی (انتقال اکسیژن به خون جنین و انتقال دی اکسید کربن به خون مادر)؛ دفعی ( محصولات متابولیک و از طریق کلیه های مادر دفع می شود؛ محافظ (از ایجاد درگیری ایمنی جلوگیری می کند، نفوذ مواد دارویی - مانع مطلق نیست)؛ غدد درون ریز (سنتز تعدادی از هورمون ها و سایر مواد فعال بیولوژیکی).
عملکرد جفت تغذیه ای است (انواع مواد مغذی، الکترولیت ها، ویتامین ها و برخی هورمون ها از بدن مادر به جنین می رسد؛ تنفسی (انتقال اکسیژن به خون جنین و انتقال دی اکسید کربن به خون مادر)؛ دفعی ( محصولات متابولیک و از طریق کلیه های مادر دفع می شود؛ محافظ (از ایجاد درگیری ایمنی جلوگیری می کند، نفوذ مواد دارویی - مانع مطلق نیست)؛ غدد درون ریز (سنتز تعدادی از هورمون ها و سایر مواد فعال بیولوژیکی).
 تبادل در سراسر سد جفت از خون مادر به جنین از خون جنین به مادر (تامین) خون (دفع) اکسیژن مواد مغذی (گلوکز، اسیدهای آمینه، اسیدهای چرب آزاد، ویتامین ها) الکترولیت ها، هورمون های استروئیدی آب (کورتیزول، استروژن) آنتی بادی ها (Ig. G) ) - ایجاد ایمنی غیرفعال جنین) پروتئین های پلاسمای خون (آلبومین) برخی هورمون های پروتئینی (تیروکسین، انسولین) گلبول های قرمز (خیلی کم) متابولیت های دی اکسید کربن (اوره، اسید اوریک، بیلی روبین، کراتین، کراتینین) هورمون های استروئیدی (گونادوتروپین کوریونی، پروژئوتروپین کوریونی، پلاسنتال) گلبول های قرمز آب (خیلی کم)
تبادل در سراسر سد جفت از خون مادر به جنین از خون جنین به مادر (تامین) خون (دفع) اکسیژن مواد مغذی (گلوکز، اسیدهای آمینه، اسیدهای چرب آزاد، ویتامین ها) الکترولیت ها، هورمون های استروئیدی آب (کورتیزول، استروژن) آنتی بادی ها (Ig. G) ) - ایجاد ایمنی غیرفعال جنین) پروتئین های پلاسمای خون (آلبومین) برخی هورمون های پروتئینی (تیروکسین، انسولین) گلبول های قرمز (خیلی کم) متابولیت های دی اکسید کربن (اوره، اسید اوریک، بیلی روبین، کراتین، کراتینین) هورمون های استروئیدی (گونادوتروپین کوریونی، پروژئوتروپین کوریونی، پلاسنتال) گلبول های قرمز آب (خیلی کم)
 انواع جفت در پستانداران اپیتلیوکوریال (خوک): پرزهای کوریونی بدون از بین بردن غشای مخاطی (غدد رحم) نفوذ می کنند. جفت دسموکوریال (مفتور): پرزهای کوریونی با بافت همبند آندومتر مادر در تماس هستند. جفت جفت جفتی (وازو-) جفت: پرزهای کوریونی با دیواره عروق آندومتر مادر بدون تخریب دیواره تماس می گیرند. جفت هموکوریال: پرزهای کوریونی اپیتلیوم، بافت همبند، دیواره عروق مادر را از بین می برند و با خونی شسته می شوند که شکاف ها را پر می کند.
انواع جفت در پستانداران اپیتلیوکوریال (خوک): پرزهای کوریونی بدون از بین بردن غشای مخاطی (غدد رحم) نفوذ می کنند. جفت دسموکوریال (مفتور): پرزهای کوریونی با بافت همبند آندومتر مادر در تماس هستند. جفت جفت جفتی (وازو-) جفت: پرزهای کوریونی با دیواره عروق آندومتر مادر بدون تخریب دیواره تماس می گیرند. جفت هموکوریال: پرزهای کوریونی اپیتلیوم، بافت همبند، دیواره عروق مادر را از بین می برند و با خونی شسته می شوند که شکاف ها را پر می کند.
لقاح
لقاحفرآیند ادغام سلول های زایای بالغ نر (اسپرم) و ماده (تخمک) نامیده می شود که در نتیجه یک زیگوت ایجاد می شود که حامل اطلاعات ژنتیکی پدر و مادر است.
فرآیند بلوغ سلول های زایای زن و مرد بسیار پیچیده است. اسپرماتوژنز در لوله های اسپرم ساز پیچیده غدد جنسی مردانه صورت می گیرد. در دوران بلوغ با تشکیل اسپرم های بالغ با توانایی لقاح به پایان می رسد. بلوغ کامل قبل از فرآیند تقسیم کاهشی انجام می شود که در نتیجه هسته اسپرم حاوی مجموعه ای از کروموزوم هاپلوئید است.
اسپرم ها دو نوع هستند: ناقل کروموزوم های جنس X و Y. هنگامی که یک اسپرم، که حامل کروموزوم X جنسی است، با یک سلول تخمک ترکیب می شود، یک جنین ماده از زیگوت به وجود می آید، زمانی که اسپرمی که دارای کروموزوم Y جنسی است، یک جنین نر بوجود می آید (تخم همیشه یک حامل کروموزوم X جنسی).
طول اسپرم بالغ 50 تا 60 میکرون است و از سر، گردن و دم تشکیل شده است. سر اسپرم که به شکل بیضی شکل است حاوی یک هسته است که توسط لایه نازکی از پروتوپلاسم احاطه شده است. دهانه رحم دارای پروتوپلاسم حاوی یک سانتروزوم اصلاح شده است که به فرآیند خرد کردن تخمک بارور شده کمک می کند. دم از پروتوپلاسم تشکیل شده و عملکردهای حرکتی را انجام می دهد. در نتیجه حرکات نوسانی قسمت دم، اسپرم ها قادر به انجام حرکات مستقل در دستگاه تناسلی زن با سرعت 2-3 میلی متر در دقیقه هستند. توانایی حرکت اسپرم ها پس از تماس آنها با ترشح وزیکول های منی و غده پروستات حاصل می شود. این مخلوط مایع منی یا اسپرم نامیده می شود. در هنگام آمیزش جنسی، به طور متوسط 3-5 میلی لیتر مایع منی در واژن زن ریخته می شود که حاوی 300-500 میلیون اسپرم است. استفاده از میکروسکوپ الکترونی ایجاد یک ساختار بسیار پیچیده از اسپرم را ممکن کرد (شکل 3.1).
مایع منی ترکیب پیچیده ای دارد و حاوی فروکتوز، پروتئین، پروتئاز، اسید فسفاتاز، اسید سیتریک و مواد فعال بیولوژیکی - پروستاگلاندین ها است. مایع منی دارای فعالیت آنتی ژنی مشخص است، در حالی که آنتی ژن ها هم در خود اسپرم و هم در قسمت مایع اسپرم یافت می شوند. اعتقاد بر این است که تعداد اجزای آنتی ژنی در مایع منی به 12 می رسد. در فرآیند لقاح، آنتی ژن های اسپرم خنثی می شوند.
رشد تخمک ها با رشد و توسعه فولیکول های اولیه واقع در لایه کورتیکال تخمدان ها همراه است. بلوغ تخم را می توان به طور مشروط به بلوغ هسته و بلوغ سیتوپلاسم تقسیم کرد. بلوغ هسته ای به عنوان مجموعه ای از فرآیندها درک می شود که هسته را از مرحله دیپلوتن I (یا مرحله وزیکول ژرمینال) به متافاز تقسیم میوز دوم منتقل می کند. بلوغ هسته شامل تکمیل میوز نمی شود، زیرا تنها نفوذ اسپرم به تخمک باعث تکمیل تقسیم کاهش دوم می شود.
سر بدن دم
شکل 3.1. ساختار اسپرم انسان (طرح میکروسکوپی الکترونی). 1 - آکروزوم؛ 2 - هسته؛ 3 - گردن؛ 4 - میتوکندری؛ 5 - رزوه های محوری.

برنج. 3.2. تخمک بالغ احاطه شده توسط سلول های کرونایی رادیاتا.
از دوران بلوغ تا یائسگی، یک زن معمولاً در هر سیکل قاعدگی یک تخمک دارد. یک تخمک مرتبه اول با جدا شدن اولین جسم قطبی به تخمک مرتبه دوم تبدیل می شود. در زمان تخمک گذاری، تخمک مرتبه دوم در مرحله متافاز تقسیم میوز دوم مسدود می شود. بلوغ تخمک ها به دلیل امکان کشت آزمایشگاهی تخمک ها که در حال حاضر در لقاح آزمایشگاهی بسیار مورد استفاده قرار می گیرد، به طور مفصل شناخته شده است.
یک تخمک بالغ از یک هسته، سیتوپلاسم، احاطه شده توسط یک غشای براق و سلول های تاج تابشی، که بقایای سلول های گرانولوزای فولیکول است، احاطه شده است (شکل 3.2). سلول جنسی زن نیز مانند نر دارای خواص آنتی ژنی است. پوسته درخشان آن به ویژه از نظر آنتی ژن های مختلف غنی است.
قبل از تجزیه و تحلیل مکانیسم های لقاح، به عنوان مثال. فرآیند ادغام تخمک با اسپرم، مسائل مربوط به انتقال گامت باید برجسته شود. اول از همه، این مربوط به حرکت اسپرم از طریق کانال های دستگاه تناسلی زنانه است. در طول انزال طبیعی، به طور متوسط حدود 100 میلیون اسپرم وارد واژن می شود که برخی از آنها دارای ناهنجاری های مورفولوژیکی یا عملکردی خاصی هستند. بخشی از اسپرمها، از جمله اسپرمهای معیوب، در واژن باقی میمانند و تحت فاگوسیتوز قرار میگیرند. همراه با اسپرم، سایر اجزای اسپرم نیز وارد واژن می شوند که نقش ویژه ای توسط پروستاگلاندین ها ایفا می شود. تحت تأثیر آنها، فعالیت انقباضی رحم و لوله های فالوپ فعال می شود که برای انتقال طبیعی گامت ها بسیار مهم است.
از واژن که در یک زن سالم دارای محیط اسیدی است (اسیدی
محیط برای فعالیت حیاتی اسپرم نامطلوب است)، دومی به سرعت وارد مخاط دهانه رحم می شود که در حین مقاربت، تحت تأثیر انقباضات ماهیچه های دهانه رحم، از کانال دهانه رحم خارج می شود. وجود یک واکنش کمی قلیایی مخاط دهانه رحم به افزایش فعالیت حرکتی اسپرم کمک می کند.
ترکیب بهینه مخاط دهانه رحم تا زمان تخمک گذاری، عمدتاً تحت تأثیر هورمون های استروژن تخمدان، تشکیل می شود. در این دوره، میسلیوم های مخاطی به شکل زنجیره های عجیب و غریب در امتداد خطوط نیروی میدان مغناطیسی زمین قرار می گیرند. اسپرماتوزواها در امتداد میسلیوم مخاطی به سمت رحم حرکت می کنند، اما فقط با ساختار خاصی از زنجیره های میسل. حرکات آشفته اسپرم در نواحی جداری دهانه رحم بارزتر است، در حالی که برخی از اسپرم ها می توانند برای مدتی در کریپت های دهانه رحم رسوب کنند، در نتیجه نوعی ذخیره اسپرم ایجاد می شود، جایی که جریان اضافی آنها از طریق دهانه رحم به سمت بالا می رود. کانال های دستگاه تناسلی می توانند در آینده ایجاد شوند.
در قسمت های بالایی دستگاه تناسلی زن، فرآیندی به نام ظرفیت اسپرم آغاز می شود - کسب توانایی بارور شدن توسط آن به دلیل تغییرات پیچیده. در نتیجه ظرفیت سازی، اسپرم ها توانایی واکنش آکروزومی را به دست می آورند. علاوه بر این، ظرفیت در تغییرات در حرکات قسمت های دم اسپرم (وجود تحرک بیش فعال) بیان می شود. مکانیسم های دقیق ظرفیت سازی هنوز به طور کامل درک نشده اند. زمان ظرفیت برای اسپرم های مختلف متفاوت است، که ظاهراً یک واکنش تطبیقی مهم برای فرآیند لقاح است. اسپرمهای دارای ظرفیت بسیار فعال هستند، اما امید به زندگی آنها کوتاهتر از اسپرمهای بدون ظرفیت است. اسپرمهای ظرفیتدار توانایی بیشتری برای نفوذ به بافتها دارند که در فرآیند لقاح تخمک بسیار مهم است.
انتقال اسپرم به داخل رحم و سپس به لوله های فالوپ عمدتاً از طریق انقباضات ماهیچه های صاف این اندام ها انجام می شود. همچنین اعتقاد بر این است که اسفنکترهای لوله رحمی نوعی توزیع کننده برای جریان اسپرم از حفره رحم به مجرای لوله های فالوپ هستند.
همراه با انقباض لوله های فالوپ که تحت تأثیرات پیچیده هورمونی (استروژن ها، آندروژن ها، اکسی توسین) و همچنین تحت تأثیر پروستاگلاندین ها قرار دارند، علاوه بر فعالیت جنبشی بالای خود، اهمیت زیادی در ارتقاء اسپرم دارد. به عواملی مانند حرکت میکروویلی های اپیتلیوم مژک دار آندوسرویکس و جریان مایع در لومن لوله فالوپ. بنابراین، حرکت اسپرم از طریق کانال های دستگاه تناسلی زنان یک فرآیند چند جزئی بسیار پیچیده است.
نتایج تحقیقات علمی مدرن حاکی از وجود یک فرآیند دو مرحله ای انتقال اسپرم از طریق لوله های فالوپ است. در مرحله اول (کوتاه)، اسپرم شروع به ورود سریع به آمپول لوله می کند. این مرحله تنها چند دقیقه طول می کشد و با فعالیت انقباضی رحم و لوله های فالوپ تنظیم می شود. این مرحله با فاز دوم طولانی تر جایگزین می شود که طی آن اسپرم ها با سرعت بسیار کمتری به محل لقاح منتقل می شوند.
در قسمت آمپولر لوله فالوپ، تعداد طبیعی اسپرم ها در مرحله انتقال طولانی به دلیل جایگزینی مداوم اسپرم ها از حفره شکم به دلیل آن دسته از سلول های زایا که در قسمت های پایینی رسوب می کنند، در سطح مشخصی حفظ می شود. دستگاه تناسلی (کریپت های دهانه رحم و غیره).
در انتقال گامت ها و لقاح، به اصطلاح گرفتن تخمک توسط بخش آمپولار لوله فالوپ اهمیت زیادی دارد. این فرآیند شامل این واقعیت است که بخش آمپولار لوله، با فیمبریای آن، تخمدان را در محلی که تخمک گذاری رخ داده است، می پوشاند. در همان زمان، حرکت یک تخمک بالغ به آمپول لوله فالوپ تا حد زیادی تسهیل می شود. مکانیسم این پدیده هنوز به طور کامل کشف نشده است.
مسئله بقای اسپرم در دستگاه تناسلی زن ضروری است. برخی از نویسندگان معتقدند که زنده ماندن اسپرم برای چندین (تا 5) روز حفظ می شود. با این حال، باید در نظر داشت که حفظ حرکت اسپرم لزوماً نشان دهنده توانایی باروری آنها نیست.
ولی در مطلوب ترین شرایط، زمانی که سلول های زایای مردانه در مخاط دهانه رحم در برابر پس زمینه محتوای بالای استروژن در بدن قرار دارند، توانایی لقاح اسپرم تا 2 روز پس از انزال در واژن ادامه می یابد. در این رابطه، عموماً پذیرفته شده است که برای رسیدن به بارداری، تناوب بهینه رابطه جنسی در دوره قبل و بعد از تخمک گذاری باید هر 2 روز یکبار باشد. با رابطه جنسی مکرر، باروری اسپرم کاهش می یابد.
قبل از لقاح، یک تخمک بالغ از همه طرف توسط یک تاج درخشان (corona radiata) احاطه شده است. پوسته براق (zona pellucida) به وضوح قابل مشاهده است. چند دقیقه پس از پارگی فولیکول (تخمکگذاری)، تخمک وارد حفره لوله فالوپ میشود. این فرآیند توسط تعدادی از عوامل تسهیل می شود: "گرفتن" لوله فالوپ توسط فیمبریا از تخمدانی که در آن تخمک گذاری رخ داده است، جهت جریان مایع فولیکولی در حین پارگی فولیکول و غیره. نقش بسیار مهمی در حمل و نقل اولیه تخمک که از تحرک مستقل محروم است. توانایی تخمک برای لقاح به طور متوسط 24 ساعت است.روش های تشخیصی مدرن (سونوگرافی، لاپاراسکوپی) نه تنها امکان مشاهده روند تخمک گذاری، بلکه همچنین ثبت این فرآیند بر روی فیلم را فراهم می کند.
تخمکی که وارد آمپول لوله فالوپ شده است به سرعت توسط تعداد زیادی اسپرم که حامل کروموزوم های جنس X و Y هستند احاطه می شود. در زیر میکروسکوپ، اسپرم های دارای کروموزوم X کمی بزرگتر از اسپرم های حامل کروموزوم Y هستند. اسپرم های اطراف تخمک شروع به نفوذ به سلول های تاج تابشی می کنند. فرآیند نفوذ به دلیل وجود تعدادی آنزیم است که هم در سر اسپرم و هم در مایع لوله ای وجود دارد (شکل 3.3).
بلافاصله پس از ادغام غشای سلول های زاینده، یک واکنش قشری تخمک رخ می دهد که بخشی جدایی ناپذیر برای اطمینان از بلوک پلی اسپرمی است. پس از این، کروموزوم های زیگوت وارد اولین تقسیم میتوزی می شوند که 24 ساعت پس از شروع لقاح رخ می دهد. هسته یک تخمک بارور شده (زیگوت) حاوی یک دیپلوئید است

برنج. 3.3. نفوذ اسپرم به تخمک در آمپول لوله فالوپ (طرح).
مجموعه جدیدی از کروموزوم ها (46). بنابراین، ارگانیسم جدید حامل اطلاعات ژنتیکی هر دو والدین است.
پس از لقاح (بعد از 24 ساعت)، برش تخم بارور شده آغاز می شود. در ابتدا، خرد کردن همزمان است. پس از گذشت 12 ساعت از شروع ظهور 2 بلاستومر، 4 بلاستومر ظاهر می شود و به همین ترتیب. تا 96 ساعت از لحظه ادغام هسته اسپرم با هسته تخمک، جنین از 16-32 بلاستومر (مرحله مورولا) تشکیل شده است. در این مرحله تخمک بارور شده (زیگوت) وارد رحم می شود.
از آنجایی که تخم مرغ خرد کننده تحرک مستقلی ندارد، حمل و نقل آن با تعامل فعالیت انقباضی لوله فالوپ (عامل اصلی)، حرکات اپیتلیوم مژگانی اندوسالپنکس و جریان مویرگی مایع در جهت تعیین می شود. انتهای آمپولار لوله فالوپ به رحم.
انتقال تخمک از طریق لوله فالوپ تحت تأثیر هورمون ها است. همانطور که می دانید پس از تخمک گذاری، یک غده درون ریز جدید به نام جسم زرد در محل ترکیدگی فولیکول تشکیل می شود. هم هورمون های پروژسترون و هم هورمون های استروژن را ترشح می کند. این دو هورمون جنسی هستند که نقش اصلی را در تضمین سینتیک لوله های فالوپ ایفا می کنند. تحت تأثیر محتوای نسبتاً کم پروژسترون و غلظت بالاتر استروژن در خون (که بلافاصله پس از تخمک گذاری اتفاق می افتد)، صدای بخش آمپولری-ایستموس لوله افزایش می یابد. در نتیجه، تخمک در آمپول باقی می ماند، جایی که بارور می شود و فرآیند تقسیم سلولی با تشکیل بلاستومرها آغاز می شود. در آینده، یک پیشروی تدریجی تخمک بارور شده در امتداد تنگه لوله فالوپ به سمت رحم وجود دارد. تحت تأثیر افزایش غلظت پروژسترون جسم زرد، عملکرد انقباضی لوله های فالوپ شخصیت پریستالتیکی پیدا می کند، در حالی که امواج انقباضات به سمت رحم هدایت می شود. محل اتصال لوله به رحم شل می شود و تخمک از لوله فالوپ وارد حفره رحم می شود.
 آندومتر
آندومتر
برنج. 3.4. انتقال تخمک بارور شده از طریق لوله فالوپ تا زمان لانه گزینی (نمودار).
1 - تخمک در آمپول لوله فالوپ. 2 - لقاح; 3-7 - مراحل مختلف تشکیل بلاستومر. 8 - مورولا; 9، 10 - بلاستوسیست؛ 11 - کاشت.
با تجزیه و تحلیل مکانیسم های تنظیم هورمونی سینتیک لوله های فالوپ، باید تاکید کرد که تنها یک نسبت معین بین غلظت پروژسترون و استروژن در خون، تعامل فیزیولوژیکی واضح بخش های مختلف لوله فالوپ که در بالا توضیح داده شد را تضمین می کند. انتقال به موقع تخمک به داخل حفره رحم. همراه با پروژسترون و استروژن، هورمون هایی مانند تستوسترون و کورتیزول نقش مهمی در اجرای سینتیک فیزیولوژیکی لوله های فالوپ در اوایل بارداری دارند. با این حال، نقش این هورمون ها هنوز به اندازه کافی بررسی نشده است.
لانه گزینی تخمک.پس از طی 4 روز از طریق لوله فالوپ، تخمک جنین در مرحله مورولا وارد رحم می شود و در آنجا به بلاستوسیست تبدیل می شود (شکل 3.4). مرحله بلاستوسیست با این واقعیت مشخص می شود که بلاستومرها دچار تغییرات خاصی می شوند. بخشی از بلاستومرها، از نظر اندازه بزرگتر، به اصطلاح را تشکیل می دهند جنین،که از آن جنین رشد می کند. بخش دیگری از سلول ها، کوچکتر و در امتداد حاشیه تخم جنین قرار دارد، یک غشای مغذی را تشکیل می دهد - تروفوبلاستدر آینده، توسعه یافته ترین قسمت تروفوبلاست به آن تبدیل می شود جفتدر حفره رحم، بلاستوسیست به محل لانه گزینی (نیداسیون) نزدیک می شود (شکل 3.5، a، b). محلی سازی کاشت الگوهای خاص خود را دارد و ظاهراً تا حد زیادی توسط ویژگی های محلی آندومتر تعیین می شود. بلاستوسیست معمولاً در دیواره قدامی یا خلفی رحم کاشته می شود.
سپس بلاستوسیست شروع به فرو رفتن در آندومتر می کند که در این زمان به دسیدو تبدیل می شود (شکل 3.5، c، d).
 |
 |
برنج. 3.5. کاشت.
الف - بلاستوسیست قبل از کاشت. ب - تماس اولیه بلاستوسیست با غشای دسیدوال رحم. ج - غوطه ور شدن بلاستوسیست در دسیدو. د - اتمام کاشت.
دسیدو یک لایه عملکردی اصلاح شده اندومتر است. در زمان لانه گزینی تخمک بارور شده، مخاط رحم در مرحله ترشح است، غدد دندانه دار و پر از ترشح هستند، سلول های استرومایی گرد شده و حاوی مقدار زیادی گلیکوژن، لیپیدها، موکوپلی ساکاریدهای خنثی، نمک ها و عناصر کمیاب هستند. آنزیم ها و مهارکننده های آنها، ایمونوگلوبولین ها و بسیاری دیگر از ترکیبات فعال بیولوژیکی که برای فعالیت حیاتی جنین ضروری هستند.
فرآیند کاشت در درجه اول با عوامل هورمونی مرتبط است. نقش اصلی متعلق به هورمون های جنسی (استروئیدی) است. در دوران بارداری، رشد و عملکرد فعال جسم زرد تخمدان، که در محل ترکیدن فولیکول ایجاد شده است، رخ می دهد. جسم زرد مقادیر زیادی پروژسترون و اندکی کمتر استروژن ترشح می کند. این هورمونهای جنسی، از طریق گیرندههای استروئیدی خاص واقع در آندومتر، تأثیر برجستهای بر دگرگونیهای ترشحی مخاط رحم و فرآیندهای جداسازی آن دارند. علاوه بر هورمون های استروئیدی، برخی هورمون های دیگر (پرولاکتین، گلوکوکورتیکوئیدها) نیز نقش خاصی در لانه گزینی دارند.
بافت برگ به عنوان نوعی اندام غدد درون ریز ارتباط مستقیمی با لانه گزینی و مراحل بعدی رشد پس از لانه گزینی جنین دارد. مشخص شده است که آندومتر یک زن یک پرولاکتین واکنشگر ایمنی تولید می کند. از روز نهم پس از تخمک گذاری شروع به ترشح می کند، در حالی که در دوران بارداری تولید پرولاکتین بافتی به طور قابل توجهی افزایش می یابد، بنابراین، در حال حاضر، برخی از محققان تمایل دارند بافت دسیدوال را نوعی اندام غدد درون ریز در نظر بگیرند.
روند کاشت، که به طور متوسط حدود 2 روز طول می کشد، نه تنها با تغییرات قابل توجهی در عناصر سلولی غدد و استرومای آندومتر، بلکه با تغییرات همودینامیک مشخص از طبیعت محلی همراه است. در نزدیکی محل کاشت بلاستوسیست، انبساط عروق خونی و تشکیل سینوسوئیدها وجود دارد که مویرگ ها و ونول های متسع هستند. این فرآیندها نقش مهمی در فرآیندهای مبادله بین ارگانیسم مادر و جنین دارند.
جنین زایی اولیه
پس از لانه گزینی، هم جنین و هم غشاهای آن به سرعت رشد می کنند. بر روی تروفوبلاست پرزهایی تشکیل می شوند که در مراحل اولیه انتوژنز به طور مساوی تمام سطح بلاستوسیست را می پوشانند. پرزهای اولیه تشکیلات بدون عروق هستند. به تدریج، لایه بیرونی تروفوبلاست مرزهای سلولی خود را از دست می دهد و به سینسیتیوم (سیمپلاست) تبدیل می شود. لایه داخلی تروفوبلاست ساختار سلولی را حفظ می کند و سیتوتروفوبلاست نامیده می شود. فرآیندهای بیشتر تمایز تروفوبلاست با جزئیات بیشتر در بخش بعدی توضیح داده شده است (به بخش 3.2 مراجعه کنید).
جنین همزمان با تروفوبلاست رشد می کند. این فرآیند به ویژه پس از کاشت فعال می شود. سلول های جنینی به دو ندول اکتوبلاستیک و اندوبلاستیک تمایز می یابند. به زودی حفره هایی در قسمت های مرکزی این گره ها ایجاد می شود که در نتیجه ندول اکتوبلاستیک به یک وزیکول اکتوبلاستیک تبدیل می شود و ان-
در مراحل اولیه رشد، تمایز مزانشیمی نیز رخ می دهد. از یک طرف، سلول های مزانشیمی به سمت حاشیه - به تروفوبلاست رانده می شوند. از سوی دیگر، تجمع سلول های مزانشیمی در اطراف کیسه های آمنیوتیک و زرده و همچنین اطراف جنین وجود دارد. در نتیجه دیواره های وزیکول ها دولایه می شوند و خود جنین به اکتو، اندو و مزودرم متمایز می شود. این سه لایه جوانه به عنوان ماده اولیه برای تشکیل بیشتر همه اندام ها و بافت های جنین و جنین عمل می کنند.
با رشد بیشتر داخل رحمی، افزایش سریع در اندازه وزیکول آمنیوتیک وجود دارد که به حفره آمنیوتیک تبدیل می شود. در حفره، تجمع سریع مایع شفاف (مایع آمنیوتیک) وجود دارد، در حالی که دیواره وزیکول آمنیوتیک به غشای پرز نزدیک می شود و در نهایت با آن ادغام می شود. جنین شروع به پیچیدن به داخل حفره آمنیون می کند و کیسه زرده آتروفی می شود.
همزمان با رشد غشاهای جنینی، یک برآمدگی به نام آلانتویس از قسمت دمی روده اولیه جنین ایجاد می شود. به گفته آلانتویس، رگهای جنین به سمت غشای پرز «رشد» میکنند و سپس در هر پرز رشد میکنند. در نتیجه کوریون آواسکولار عروقی می شود.
پس از اتمام مراحل اولیه انتوژنز، جنین توسط مایع آمنیوتیک و سه غشای دسیدوآ، پرز و آبی احاطه می شود.
دسیدو یک لایه عملکردی آندومتر است که به دلیل بارداری تغییر شکل داده است. تا زمان لانه گزینی، آندومتر در فاز ترشحی است. از دو لایه تشکیل شده است: فشرده و اسفنجی. لایه فشرده، رو به حفره رحم، حاوی مجاری دفعی غدد رحم و سلول های استرومای آندومتر است که در دوران بارداری به دسیدوال تبدیل می شوند. لایه اسفنجی عمدتاً از غدد تشکیل شده است.
در طول رشد تخمک جنین، decidua parietalis و decidua capsularis کشیده شده، نازک تر شده و به یکدیگر نزدیک می شوند. در ماه 4-5 بارداری، تخمک جنین در حال حاضر کل حفره رحم را اشغال می کند و decidua parietalis با decidua capsularis ادغام می شود. برعکس، decidua basalis به شدت هیپرتروفی می شود و به قسمت مادری جفت تبدیل می شود. در این قسمت از جفت پرزهای کوریونی متعددی وجود دارد که فضای بین پرزهای اطراف آنها تشکیل شده است.
غشای decidual یک لایه مغذی و محافظ برای جنین است: عملکرد تغذیه ای آن عمدتاً در مراحل اولیه رشد داخل رحمی انجام می شود ، نقش محافظتی به طور کامل با فعالیت فاگوسیتیک بالا آشکار می شود. لیز بافت دسیدوال
میکروارگانیسم ها و سموم آنها را غیر فعال می کند، همچنین در سنتز کربوهیدرات ها، لیپیدها و پروتئین ها شرکت می کند. پرولاکتین و پروستاگلاندین را سنتز می کند. بنابراین، دسیدوا نقش بسیار مهمی در لانه گزینی و رشد بیشتر جنین و جنین دارد.
آمنیون (غشای آبی) رو به جنین است. جفت را می پوشاند و به بند ناف می رسد و در ناحیه حلقه ناف با پوست جنین ادغام می شود. از نظر ماکروسکوپی، آمنیون یک غشای شفاف نازک است. در طول جنین زایی، آمنیون از وزیکول اکتوبلاستیک ایجاد می شود. اپیتلیوم آمنیون از اکتودرم و پایه بافت همبند از مزودرم تشکیل شده است.
در مراحل اولیه رشد، اپیتلیوم آمنیون توسط سلول های چند ضلعی بزرگ نشان داده می شود، از ماه 3 بارداری به مکعب تبدیل می شود. سلول های اپیتلیال آمنیون حاوی لیپیدها، پلی ساکاریدها،
پروتئین ها، ترکیبات فسفر، و همچنین تعدادی از آنزیم های دخیل در فرآیندهای متابولیسم و تبادل هورمون های استروئیدی.
آمنیون، همراه با کوریون صاف، نقش فعالی در تبادل مایع آمنیوتیک و همچنین در تبادل پارا جفتی دارد. با توجه به خواص فیزیکی، غشاهای جنین با یکدیگر متفاوت هستند. از آنجایی که غشای آمنیوتیک بسیار متراکم است و فشار چندین برابر بیشتر از کوریون صاف را تحمل می کند، پارگی کوریون صاف زودتر از آمنیون در زایمان اتفاق می افتد.
جفت
جفتانسان ساختاری از نوع هموکوریال دارد. این نوع جفت با وجود تماس مستقیم خون مادر با کوریون به دلیل نقض یکپارچگی دسیدوای رحم با باز شدن عروق آن مشخص می شود.
ظهور جفت هموکوریال در تکامل بالاترین مرحله است. این نشان دهنده پیچیده ترین رابطه بین سیستم های عملکردی مادر و جنین است.
رشد و عملکرد جفت
قسمت اصلی جفت پرزهای کوریونی است - مشتقات تروفوبلاست. در مراحل اولیه انتوژنز، تروفوبلاست برآمدگی های پروتوپلاسمی متشکل از سلول های سیتوتروفوبلاست (پرزهای اولیه) را تشکیل می دهد. پرزهای اولیه فاقد عروق هستند و تامین مواد مغذی و اکسیژن به بدن جنین از خون مادری که آنها را احاطه کرده است، طبق قوانین نسبتاً ساده اسمز و انتشار انجام می شود. در پایان هفته دوم بارداری، بافت همبند به شکل پرزهای اولیه و پرزهای ثانویه رشد می کند. آنها بر اساس بافت همبند هستند و پوشش بیرونی توسط اپیتلیوم (تروفوبلاست) نشان داده می شود. پرزهای اولیه و ثانویه هر دو به طور مساوی روی سطح تخمک توزیع می شوند.
اپیتلیوم پرزهای ثانویه از دو لایه تشکیل شده است: لایه سیتوتروفوبلاست (لایه Langhans) و سینسیتیوم (سیمپلاست). لایه سیتوتروفوبلاست از سلول های گرد با سیتوپلاسم سبک تشکیل شده است. هسته های سلولی بزرگ هستند. در سینسیتیوم، مرزهای سلولی عملاً قابل تشخیص نیستند، سیتوپلاسم تیره، دانه ای، با حاشیه قلم مو است. هسته ها نسبتا کوچک، کروی یا بیضی شکل هستند.
از هفته سوم رشد جنینی، فرآیند بسیار مهمی از رشد جفت آغاز می شود که شامل عروقی شدن پرزها و تبدیل آنها به عروق ثالثیه است. تشکیل رگ های جفتی هم از آنژیوبلاست های جنین و هم از عروق نافی که از آلانتویس رشد می کنند اتفاق می افتد.
عروق آلانتوئیس به پرزهای ثانویه تبدیل می شوند که در نتیجه هر پرز ثانویه عروقی می شود. به این ترتیب مهمترین فرآیند رشد داخل رحمی - عروقی شدن کوریون انجام می شود. برقراری گردش خون آلانتوئید تبادل شدید بین ارگانیسم های جنین و مادر را فراهم می کند.
▲ تبدیل پرزهای ثانویه به پرزهای ثالثی مهمترین دوره بحرانی رشد جنینی محسوب می شود، زیرا آلانتویس حساسیت بسیار بالایی به عملکرد عوامل مخرب محیطی دارد. آسیب آن با مرگ رگ های خونی همراه است که در نتیجه مهمترین فرآیند متوقف می شود- عروقی شدن کوریون و این به نوبه خود منجر به مرگ جنین در مراحل اولیه می شود.
در مراحل اولیه رشد داخل رحمی، پرزهای کوریونی به طور مساوی تمام سطح تخمک جنین را می پوشانند. با این حال، از ماه دوم انتوژنز، در سطح بزرگتر تخم جنین، پرزها آتروفی میشوند، در حالی که در همان زمان، پرزهای رو به قسمت پایه دسیدوآ به شکلی مجلل رشد میکنند. به این ترتیب یک کوریون صاف و منشعب تشکیل می شود.
توسعه و تمایز بیشتر کوریون با نکات اصلی زیر مشخص می شود. در سن حاملگی 6-5 هفته، ضخامت سین سیتیوتروفوبلاست از ضخامت لایه لانگانس بیشتر می شود و از 9 تا 10 هفتگی، سینسیتیوتروفوبلاست به تدریج نازک می شود و تعداد هسته های آن افزایش می یابد. در سطح آزاد سینسیتیوتروفوبلاست رو به فضای بین پرزها، برآمدگی های سیتوپلاسمی نازک طولانی (میکروویلی) به وضوح قابل مشاهده است که سطح جذب جفت را به میزان قابل توجهی افزایش می دهد. در ابتدای سه ماهه دوم بارداری، تبدیل شدید سیتوتروفوبلاست به سینسیتیوم رخ می دهد که در نتیجه لایه Langhans در بسیاری از مناطق به طور کامل ناپدید می شود.
در پایان بارداری، فرآیندهای دگرگونی-دیستروفی در جفت شروع می شود که گاهی اوقات پیری جفت نامیده می شود. از خونی که در فضای بین پرزها در گردش است، فیبرین (فیبرینوئید) شروع به ریزش می کند که عمدتاً در سطح پرزها رسوب می کند. از دست دادن این ماده به فرآیندهای تشکیل میکرو ترومبوز و مرگ بخش های فردی از پوشش اپیتلیال پرزها کمک می کند. پرزهای پوشیده شده با فیبرینوئید تا حد زیادی از تبادل فعال بین مادر و جنین حذف می شوند.
نازک شدن آشکار غشای جفت از 25 میکرون در ابتدای بارداری تا 5 میکرون در انتهای آن وجود دارد. استرومای پرزها فیبری تر و همگن تر می شود. اندوتلیوم مویرگی کمی ضخیم می شود. نمک های آهک اغلب در مناطق دیستروفی رسوب می کنند. همه این تغییرات در عملکردهای اصلی جفت (تنفسی، تغذیه ای، متابولیک، غدد درون ریز و غیره) منعکس می شود.
همراه با فرآیندهای فرورفتگی در جفت در دوران بارداری، پدیده هایی با طبیعت مخالف نیز مشاهده می شود. افزایش پرزهای جوان وجود دارد که تا حد زیادی عملکرد پرزهای از دست رفته را جبران می کند. با این حال، رشد پرزهای جوان فقط تا حدی عملکرد جفت را به عنوان یک کل بهبود می بخشد. در نتیجه در پایان بارداری، عملکرد جفت کاهش می یابد.
ساختار جفت بالغ (شکل 3.8). جفت بالغ از نظر ماکروسکوپی بسیار شبیه یک کیک نرم ضخیم است. جرم جفت 500-600 گرم قطر 15-18 سانتی متر و ضخامت 3-2 سانتی متر جفت دارای دو سطح است: سطح مادری رو به دیواره رحم و سطح میوه ای به سمت دیواره رحم. جنین

برنج. 3.8. ظاهر جفت بالغ، یک - قسمت مادری؛ ب - قسمت میوه.
سطح مادرجفت رنگ قرمز مایل به خاکستری دارد و بقایای قسمت پایه دسیدواست.
سطح میوهاز بالا با یک غشای آمنیوتیک براق پوشیده شده است که در زیر آن عروقی که از محل اتصال بند ناف به حاشیه جفت می آیند به کوریون نزدیک می شوند. قسمت اصلی جفت جنین توسط پرزهای کوریونیک متعددی نشان داده می شود که به شکل های لوبدار - لپه ها یا لوبول ها ترکیب می شوند. تعداد آنها به 15-20 می رسد. لوبول های جفت در نتیجه جدا شدن پرزهای کوریونی توسط پارتیشن ها (سپتا) که از صفحه پایه بیرون می آیند تشکیل می شوند. هر کدام از این لوبول ها رگ بزرگ خود را دارند.
ساختار میکروسکوپی پرز بالغ مرسوم است که بین دو نوع پرز تشخیص داده شود: آزاد و ثابت (لنگر). پرزهای آزاد و بیشتر آنها در فضای بین پرزهای دسیدو فرو رفته و در خون مادر شناور هستند. در مقابل، پرزهای لنگر به دسیدووا بازالیس متصل هستند و جفت را به دیواره رحم ثابت می کنند. در مرحله سوم زایمان، اتصال این گونه پرزها با دسیدو قطع می شود و تحت تأثیر انقباضات رحمی، جفت از دیواره رحم جدا می شود.
هنگام مطالعه میکروسکوپی ساختار یک پرز بالغ، می توان تشکیلات زیر را متمایز کرد (شکل 3.9):
سینسیتیوم بدون مرزهای سلولی واضح؛
لایه (یا بقایای) سیتوتروفوبلاست.
پرزهای استروما؛
اندوتلیوم مویرگ که در لومن آن عناصر به وضوح قابل مشاهده است
خون جنین
میکروسکوپ الکترونی پرزهای کوریونی نشان داد که سینسیتیوم دارای میکروویل های متعددی در سطح خود است که سطح تبادل جفت را به میزان قابل توجهی افزایش می دهد.
گردش خون رحمی جفتی. اگر برنامه ای وجود دارد
جریان خون در رحم با کمک 150 تا 200 شریان مارپیچ مادری انجام می شود که به یک فضای بین پرزهای وسیع باز می شوند. شریان های مارپیچی ساختاری عجیب دارند، دیواره های آنها فاقد لایه عضلانی است و دهان ها قادر به انقباض و انبساط نیستند.
شریان های مارپیچی مقاومت عروقی کمی در برابر جریان خون دارند. بر خلاف شریان های رحمی، که در آنها کاهش شدید مقاومت عروقی از هفته 12-13 بارداری مشاهده می شود، در شریان های مارپیچی، همانطور که با استفاده از داپلر ایجاد شد، این روند از هفته 6 بارداری انجام می شود. بارزترین کاهش مقاومت عروقی در شریان های مارپیچی در هفته 13-14 بارداری مشاهده می شود که از نظر مورفولوژیکی نشان دهنده تکمیل تهاجم پرزهای تروفوبلاست به دسیدواست.
ویژگی های توصیف شده همودینامیک در اجرای انتقال بی وقفه خون شریانی از بدن مادر به جنین اهمیت زیادی دارد. خون شریانی خروجی پرزهای کوریونی را میشوید و در عین حال اکسیژن، مواد مغذی لازم، بسیاری از هورمونها، ویتامینها، الکترولیتها و سایر مواد شیمیایی و همچنین ریز عناصر لازم برای رشد و نمو مناسب جنین را به خون جنین میرساند. خون حاوی CO 2 و سایر محصولات متابولیکی جنین در منافذ وریدی وریدهای مادر ریخته می شود که تعداد کل آنها بیش از 180 است.
جریان خون در فضای بین پرز در پایان بارداری بسیار شدید است و به طور متوسط 500-700 میلی لیتر خون در دقیقه است.
ویژگی های گردش خون در سیستم مادر-جفت-جنینرگ های شریانی جفت پس از خروج از بند ناف به صورت شعاعی بر حسب تعداد لوبول های جفت (لپه ها) تقسیم می شوند. در نتیجه انشعاب بیشتر عروق شریانی، شبکه ای از مویرگ ها در پرزهای انتهایی تشکیل می شود که خون از آن در سیستم وریدی جمع آوری می شود. وریدهایی که خون شریانی در آنها جریان دارد در تنه های وریدی بزرگتر جمع آوری می شوند و در نهایت به داخل سیاهرگ بند ناف می ریزند (شکل 3.10).
 |
برنج. 3.10. گردش خون در سیستم مادر - جفت - جنین.
1 - میومتر; 2 - پرزهای آزاد; 3 - شمع لنگر؛ 4 - دسیدوا; 5 - فضای بین پرز; 6 - شریان های مارپیچی; 7 - صفحه کوریونی؛ 8 - اپیتلیوم کوریونی؛ 9 - اپیتلیوم آمنیوتیک؛ 10 - بند ناف; 11 - ورید بند ناف; 12 - شریان های بند ناف; 13 - رسوب فیبرینوئید.
گردش خون در جفت توسط انقباضات قلبی مادر و جنین پشتیبانی می شود. نقش مهمی در ثبات این گردش خون نیز به مکانیسم های خود تنظیمی گردش خون رحمی جفتی تعلق دارد.
وظایف اصلی جفت. قرار دادن
جنین شناسی.
جنین شناسی شماره 1
مفهوم پیشزایی و جنینزایی. دوره ها و مراحل اصلی جنین زایی در انسان. سلول های جنسی انسان، ویژگی های ساختاری و ژنتیکی آنها.
جنین زایی -
پروژنز -دوره رشد و بلوغ سلول های زایا - تخم ها و اسپرم ها، در نتیجه، مجموعه هاپلوئیدی از کروموزوم ها در سلول های زایای بالغ ظاهر می شود، ساختارهایی تشکیل می شود که توانایی آنها را برای لقاح و توسعه یک ارگانیسم جدید تضمین می کند.
سلول های جنسی - علائم:
مجموعه کروموزوم هاپلوئید
نسبت هسته ای-سیتوپلاسمی تغییر یافته - نسبت حجم هسته و سیتوپلاسم
متابولیسم سلولی تغییر کرد
سلول ها به شدت متمایز هستند (قابلیت تقسیم ندارند)
تخم مرغ (توسط بهر کشف شد ) - دارای اولمما، اوپلاسم (سیتوپلاسم)، هسته؛ همه اندامک ها به جز سانتریول ها رشد می کنند؛ زرده در میان آخال ها غالب است.
خصوصیات فیزیولوژیکی: هسته نه از نظر رونویسی و نه از نظر همانندسازی غیر فعال است، یعنی. منفعلانه رفتار می کند؛ تخم مرغ آنزیم ها، فاکتورها و گلیکوژن را جمع می کند.
ویژگی های ساختاری: دارای لایه قشر سیتوپلاسم- هیالوپلاسم محیطی با گرانول های قشری (موکوپلی ساکاریدها، پروتئین ها، آنزیم ها)؛ قطبی- دو قطب متمایز می شود: حیوانی (ارگانل ها متمرکز هستند) و رویشی (شامل< или >مقدار پروتئین)
در انسان زرده کوچک، اولیگولسیتال ثانویه، ایزولسیتال
تخم مرغ توسط یک غشای شفاف (لسیتال) احاطه شده است - زونا پلوسیدا (از پروتئین های Zp تشکیل می شود). در میان آنها پروتئین Zp2 وجود دارد - از پلی اسپرمی جلوگیری می کند، گیرنده Zp3 برای اسپرم.
تخمک توسط سلول های فولیکولی احاطه شده است که تغذیه را به آن می رسانند - آنها یک تاج درخشان را تشکیل می دهند.
اسپرماتوزوئید -چهار بخش متمایز می شوند: سر (شامل یک هسته بزرگ و یک آکروزوم - یک لیزوزوم اصلاح شده)، گردن (سانتریول پروگزیمال)، بدن (مارپیچ میتوکندری و سانتریول دیستال)، دم (که توسط یک تاژک نشان داده می شود)
حداکثر توانایی کود دهی تا دو روز.
مهاجرت مستقیم اسپرم توسط کموتاکسی تعیین می شود و رئوتاکسی، pH و مخاط از شاخص های مهم هستند. کاپوسیتوز رخ می دهد - تحت تأثیر اسرار دستگاه تناسلی زن، اسپرم توانایی لقاح را به دست می آورد.
ترویج اسپرم توسط پروستاگلاندین ها (بر روی پوشش لوله های فالوپ عمل می کند) تسهیل می شود.
EMRIOLOGY №2
مراحل اصلی جنین زایی مفهوم لقاح ویژگی های لقاح در انسان: مورفولوژی، شرایط لازم. مفهوم زیگوت.
جنین زایی -رشد جنین انسان 280 روز طول می کشد، به سه دوره تقسیم می شود: اولیه (هفته اول رشد)، جنینی (2-8 هفته رشد - تخمگذاری اندام های اصلی)، جنینی (9 هفته - قبل از تولد).
جنین زایی اولیه به مراحل زیر تقسیم می شود:
ZYGOTE - آغاز سنتز DNA و پروتئین
تکه تکه شدن - آغاز سنتز انواع اصلی RNA
مورولا - سلول های جنین همه توان (قابل تعویض) هستند.
BLASTOCYSTA - از دست دادن تمام توان وجود دارد و سلول ها برای تشکیل ساختارهای ژرمینال و خارج جنینی تعیین می شوند.
GASTRULA - لایه های زاینده و سلول های بنیادی ظاهر می شوند
اندام زایی - اندام ها از بافت تشکیل می شوند، ابتدایی اندام ها از کلون های سلولی تشکیل می شوند.
لقاح -ادغام سلول های زایای نر و ماده که در نتیجه آن مجموعه دیپلوئید کروموزوم های مشخصه این گونه بازسازی می شود و یک سلول کیفی جدید ظاهر می شود - یک زیگوت (تخم مرغ بارور شده یا جنین تک سلولی)
سه مرحله:
روتاکسیس مثبت - حرکت اسپرم در برابر جریان مایع ترشح شده از لوله های فالوپ
کموتکسیس - سلول های جنسی هورمون ها (گامون) ترشح می کنند. ♀ - gynogamones، ♂ - androgamones.
CAPOCYTATION - کسب توانایی لقاح توسط اسپرم تحت اثر ترشح لوله های فالوپ
فعال سازی تخمک - قسمتی از غشای تخمک که از اسپرم به دست می آید برای یون های Na نفوذپذیر است ← تغییرات قدرت سلولی ← یون های کلسیم از انبار سلولی به داخل سیتوپلاسم خارج می شوند ← اگزوسیتوز گرانول های قشر مغز. این منجر به تشکیل یک غشای لقاح می شود - تغییر در خواص زونا پلوسیدا، از پلی اسپرمی جلوگیری می کند.
اسپرم در داخل تخمک - 12 ساعت دوام می آورد و در نتیجه یک زیگوت تشکیل می شود. در این مرحله، هسته سلول های زاینده نامیده می شوند پیش هستهماتریکس هسته ای آنها شل می شود، پوسته ها ناپدید می شوند - مرحله سینکاریون. پرونوکلئوزها به یکدیگر نزدیک می شوند، در هر یک از آنها دو برابر شدن DNA رخ می دهد و کروموزوم هایی تشکیل می شود که مخلوط شده و در صفحه متافاز اولین تقسیم میوز قرار می گیرند.
تعامل از راه دور
فعل و انفعالات تماسی - اسپرم تاج درخشنده را هل می دهد و کانالی را در زونا شفاف تشکیل می دهد. اسپرم با گیرنده (پروتئین Zp3) تماس می گیرد و یک واکنش آکروزومی آغاز می شود - اگزوسیتوز محتویات آکروزوم برای تخریب موضعی غشای شفاف. در داخل سر رخ می دهد: یون های Ca و Na به داخل، H2 به بیرون. نتیجه - افزایش غلظت کلسیم باعث ایجاد یک سری فرآیندها می شود که منجر به افزایش pH داخل سلولی می شود و این باعث واکنش آکروزومی می شود. نتیجه واکنش آکروزومی تشکیل کانالی در غشای شفاف است که اسپرم از آن عبور می کند. غشای اسپرم در اولمما تعبیه شده است.
مرحله سوم
ارزش اسپرم -½ کروموزوم در زیگوت پدری هستند. ژنوم میتوکندری پدر؛ یک پروتئین سیگنال برش را معرفی می کند. بلوک میوز برداشته می شود. توسط ساختار ژنتیکی ارگانیسم تعیین می شود.
پیامد لقاح، تغییر در حجم زیگوت، دپلاریزاسیون غشای پلاسمایی و تشکیل غشای لقاح است.
جنین شناسی شماره 3
مفهوم تکه تکه شدن جنین. خصوصیات له کردن انسان: انواع خرد کردن، زمان جنین زایی، مدت زمان، شرایط. ساختار جنین در مرحله لانه گزینی در انسان.
تقسیم کردن -تقسیم متوالی میتوزی زیگوت به سلول ها (بلاستومرها) بدون رشد سلول های دختر به اندازه مادر.
سلول های تقسیم شده رشد نمی کنند، واگرا نمی شوند و دیپلوئید باقی می مانند. چرخه x میتوزی به دلیل دوره های G1 و G2 کاهش می یابد.
جوهر خرد کردن: یک ارگانیسم چند سلولی تشکیل می شود، روابط هسته ای-سیتوپلاسمی بازسازی می شود.
در انسان تکه تکه شدن: کامل(مواد زیگوت کاملا خرد شده است) نامتقارن(افزایش تعداد بلاستومرها با نقض پیشرفت هندسی رخ می دهد.
انواع بلاستومرها: روشن کوچک و تیره بزرگ
پس از 30 ساعت، اولین شیار شکافی عبور می کند (دو بلاستومر تشکیل می شود)
بعد از 40 ساعت - 4 بلاستومر
بعد از 50-60 - مورولا (توت) تشکیل می شود
سبک های کوچک - در امتداد حاشیه، یک ترفوبلاست را تشکیل می دهند
تاریک های بزرگ - در داخل، یک جنین را تشکیل می دهند.
در روز پنجم، بلاستولا (بلاستوسیست) تشکیل می شود. ویژگی آن بلاستوکول است.
در ابتدا ندول ژرمینال از رویانوبلاست و سپس سپر ژرمینال تشکیل می شود.
در روز پنجم، بلاستوسیست وارد رحم می شود.
در روز 6-7 مرحله اول گاسترولاسیون اتفاق می افتد و لانه گزینی اتفاق می افتد.
لانه گزینی - ورود جنین به مخاط رحم، دو مرحله:
ACCESSION (چسبندگی) - سلول های تروفوبلاست به طور فعال شروع به تقسیم و ادغام می کنند تا یک سمپلاستوتروفوبلاست - یک پرز کوریونی را تشکیل دهند.
نفوذ (تهاجم) - سمپلاستوتروفوبلاست آنزیم های پروتولیتیک ترشح می کند که به طور متوالی اپیتلیوم، بافت همبند و رگ های خونی مخاط رحم را تخریب می کند.
در ابتدا، جنین از بافت های تخریب شده مادر - یک نوع تغذیه هیستوتروفیک - تغذیه می کند و سپس از خون مادر - یک نوع تغذیه هماتوتروفیک تغذیه می کند.
جنین شناسی شماره 4
مفهوم و مکانیسم های اساسی گاسترولاسیون. انواع گاسترولاسیون. خصوصیات مورفولوژیکی و زمانی گاسترولاسیون در انسان ساختار جنین انسان دو هفته ای. ایده هایی در مورد دوره های بحرانی توسعه
گاسترولاسیون -یک فرآیند پیچیده از تغییرات شیمیایی و مورفوژنتیک، همراه با تولید مثل، رشد، حرکت هدایت شده و تمایز سلول ها، که منجر به تشکیل لایه های جوانه می شود: بیرونی - اکتودرم، داخلی - آندودرم، میانی - مزودرم. اینها منابع اولیه بافت ها و اندام ها، مجموعه اندام های محوری هستند.
روشهای گاسترولاسیون:
INVAGINATION (invagination) - قسمت پایین تا سقف کشیده می شود
EPIBOLIA (گرفتگی) - مشخصه سلول های بزرگ پر از زرده.
مهاجرت (اخراج)
لایه برداری (شکاف)
در انسان در دو مرحله انجام می شود:
لایه برداری - سپر ژرمینال (جنین بلاست) به تقسیم می شود اپی بلاست(همه ابتدایی ها به جز اندودرم) و هیپوبلاست(اندودرم).
مهاجرت (در روزهای 14-17) - حرکت توده های سلولی از سر و دم به پایان می رسد، سپس آنها به صورت موازی پیش می روند و یک نوار اولیه را تشکیل می دهند. موادی که در سر انتها خاموش بود شروع به حرکت می کند و به سمت جریان اول می رود. مواد سلولی دوباره مرتب شده و یک گره اولیه با یک گودال در مرکز تشکیل می شود. پایه پیشکوردال از طریق گره اولیه (هنسن) حرکت میکند و به داخل اندودرم (تا انتهای سر جنین) میرود. پایه وتر از طریق گره اولیه بین اکتو و اندودرم قرار دارد و یک وتر را تشکیل می دهد. مزودرم روی لبه های نوار اولیه با دو بال جمع می شود و به سمت داخل می رود که در طرفین نوتوکورد قرار دارد. اکتودرم همیشه در یک لایه حرکت می کند، مزودرم حرکاتی شبیه آمیب انجام می دهد، اندودرم نسبتاً بی حرکت است.
عوامل ایجاد کننده گاسترولاسیون:
رشد ناهموار در نواحی مختلف جنین
مکانیکی (بلاستومرها به راحتی تغییر شکل می دهند)
شدت جذب آب متفاوت است
شکل پذیری متفاوت غشای سلولی
توانایی سلول ها برای فاگوسیتوز
فعل و انفعالات بین سلولی
جنین شناسی شماره 5.
مفهوم تمایز لایه های جوانه. مفهوم استقرا به عنوان عاملی که باعث تمایز می شود. تمایز لایه های جوانه و تشکیل ابتدایی بافت ها و اندام ها در جنین انسان.
تمایز اکتودرم اولیه:
لوله عصبی(نوروسیت ها و نوروگلیا شبکیه و اندام بویایی، نوروسیت ها و نوروگلیا مغز و نخاع)
تاج عصبی و صفحات گانگلیونی(نوروسیت ها و نوروگلیاهای ستون فقرات و گانگلیون های اتونوم، بافت کرومافین و بصل الکلیوی آدرنال)
پلاکدها(عناصر اپیتلیال گوش داخلی)
اکتودرم پوست(اپیدرم و مشتقات آن، اپیتلیوم قرنیه، اپیتلیوم حفره دهان و مشتقات آن، اپیتلیوم مقعدی رکتوم، مینا و کوتیکول دندان، پوشش اپیتلیال واژن)
بشقاب پیش کوردال(اپیتلیوم حفره دهان و مری، اپیتلیوم نای، برونش و ریه)
اکتودرم ژرمینال
اکتودرم خارج جنینی(اپیتلیوم آمنیون و بند ناف)
تمایز اندودرم های اولیه
آندودرم لوله روده(اپیتلیوم روده، معده، کبد، پانکراس)
آندودرم ویتلین(اپیتلیوم آلانتویس و محل زرده)
اندودرم ژرمینال
اندودرم خارج جنینی
تمایز مزودرم
سومیت ها
میوتوم(بافت ماهیچه اسکلتی)
اسکلروتوم(بافت غضروفی و استخوانی)
درماتوم(پایه بافت همبند پوست)
مزانشیم(بافت همبند اپیتلیوم اگزوسلومیک، آمنیون و کوریون کیسه زرده)
Splanchnotome
صفحات جدایی و احشایی
مزودرم ژرمینال
ساقه سگمنتال نفروگونوتوما(اپیتلیوم غدد جنسی و مجرای دفران و کلیه ها)
مجرای پارامزونفریک(پوشش پوششی واژن، رحم، مجرای تخمک)
Splanchnotome(بافت ماهیچه ای مخطط قلب، قشر آدرنال، مزوتلیوم)
مزانشیم(میکروگلیا، بافت ماهیچه صاف، عروق خونی، بافت همبند، سلول های خونی و اندام های خونساز)
مزودرم خارج جنینی
سلف ها -در نواحی خاصی از جنین بوجود می آیند و باعث رشد نواحی دیگر در جهت خاصی می شوند.
جنین شناسی شماره 6.
مفهوم و اهمیت اندام های خارج جنینی. ظهور آنها در تکامل. اندام های خارج جنینی در انسان آموزش، ساختار، معنی آمنیون، کیسه زرده، آلانتوا. چین تنه، تشکیل آن، نقش.
اندام های جواهر اضافی (موقت، موقت) -در فرآیند جنین زایی در خارج از بدن جنین ایجاد می شود، عملکردهای مختلفی را انجام می دهد که رشد و نمو خود جنین را تضمین می کند.
برخی از این اندام های احاطه کننده جنین را غشای ژرمینال می نامند. این اندام ها عبارتند از: آمنیون، کیسه زرده، آلانتوئیس، کوریون، جفت
در انسان، این اندام ها زود گذاشته می شوند. در روز یازدهم رشد، تخلیه مزودرم شروع می شود، که حفره بلاستوسیست را پر می کند. مزودرم خارج جنینی از اپی بلاست خارج می شود که در جلوی اپی بلاست رشد می کند و وزیکول آمنیوتیک آینده را تشکیل می دهد. سپس اکتودرم خارج جنینی در امتداد آن رشد می کند. مزانشیم از اپی بلاست خارج می شود که در جلوی هیپوبلاست رشد می کند و کیسه زرده آینده را تشکیل می دهد. بعداً آندودرم خارج جنینی در امتداد آن رشد می کند و کیسه زرده تشکیل می شود.
آلانتویز -در روز پانزدهم جنین زایی به صورت بیرون زدگی دیواره لوله روده ایجاد می شود. رگ های خونی را به پرزهای کوریون هدایت می کند و با کاهش، بخشی از بند ناف می شود.
کوریون -در پایان هفته دوم جنین زایی، تروفوبلاست شروع به تشکیل پرزهای کوریونی اولیه می کند. در آغاز هفته سوم، مزانشیم به آنها رشد می کند و پرزهای کوریونی ثانویه تشکیل می شوند. به زودی، مزانشیم به بافت همبند و عروق خونی تمایز می یابد، بنابراین پرزهای کوریونی ثانویه تشکیل می شود که قسمت جنینی جفت را تشکیل می دهد. پرزهای کوریون، در مجاورت غشای سقوط اصلی، به شدت رشد می کنند و یک کوریون شاخه ای تشکیل می شود که با اتصال به غشای اصلی سقوط، جفت را تشکیل می دهد.
YOLK SAC -تا 7-8 هفته رشد در خونسازی شرکت می کند. سلول های زایای اولیه در اینجا تشکیل می شوند، بعداً تکامل می یابند و به شکل یک لوله باریک در بند ناف یافت می شوند.
آمنیون -به توسعه بزرگ دست می یابد، یک محیط آبی مطلوب برای رشد جنین ایجاد می کند
???????????????????????????????????????????????????????????????????????????????????????????????????
جنین شناسی شماره 7
جفت. معنای آن، ظهور در تکامل. انواع جفت. جفت انسان: نوع، ساختار، عملکردها. ساختار و اهمیت سد جفتی.
پلاسنتا -پرزهای کوریونی در مجاورت غشای سقوط اصلی به شدت رشد می کنند و کوریون شاخه ای تشکیل می شود که به غشای سقوط اصلی متصل می شود و جفت یا محل کودک را تشکیل می دهد.
جفت بالغ قطر 17-20 سانتی متر ضخامت 2-4 سانتی متر وزن 500-600 گرم.
ساختار دیسکوئیدی، هموکوریال، پرزدار است.
انواع جفت:
در ارتباط با ویژگی های نفوذ پرزهای کوریونی به رحم، آنها به موارد زیر تقسیم می شوند:
اپیتلیال کوریونیک - پرزهای کوریونی اپیتلیوم رحم (اسب، خوک) را از بین نمی برند.
دسموکوریال - پرزهای کوریونی اپیتلیوم رحم را از بین می برند و با بافت همبند (نشخوارکنندگان و آرتیوداکتیل ها) در تماس هستند.
اندوتلیوکوریال - پرزهای کوریونی اپیتلیوم رحم، بافت همبند را از بین می برند و با اندوتلیوم عروقی (شکارچیان و لاستونژیا) تماس پیدا می کنند.
هموکوریال - پرزها اپیتلیوم رحم، بافت همبند و دیواره عروق را تخریب می کنند و مستقیماً با خون مادر (پریمات، انسان) تماس می گیرند.
بر اساس ماهیت تغذیه:
کوریون پروتئین ها را از بدن مادر جذب می کند، آنها را به اسیدهای آمینه تجزیه می کند. سنتز پروتئین های اختصاصی جنین در کبد جنین اتفاق می افتد. اینها شامل جفت اپیتلیوکوریال و دسموکوریال است. پس از تولد، توله ها قادر به تغذیه و حرکت مستقل هستند.
کوریون اسیدهای آمینه را جذب می کند، پروتئین های اختصاصی جنین (همه جفت های دیگر) را سنتز می کند، توله ها پس از تولد برای مدت نسبتا طولانی سازگار می شوند.
ساختار جفت:
قسمت جنینی
غشای آمنیوتیک - از یک لایه اپیتلیوم منشوری و بافت همبند خارج جنینی تشکیل شده است.
بشقاب کریال
ویلاهای کریال
استروما (پایه) صفحه کوریونی و پرزها بافت همبند سست (شامل یک ماده بی شکل، فیبرهای آرژیروفتال، بسیاری از مویرگ ها و سلول های کاشچنکو-هوبائر - ماکروفاژهای اولیه) است. از بالا، صفحات با اپیتلیوم تروفوبلاست پوشیده شده است. در سه ماهه اول با سیتوتروفوبلاست، در سه ماهه دوم با سیتو- و سمپلاستوتروفوبلاست پوشیده می شوند. در سوم - فیبرینوئید (توده اکسی دوست همگن که محصول سرم خون و تجزیه عناصر تروفوبلاستیک است). در دوران بارداری، پرزها به شدت افزایش یافته و شاخه های آنها افزایش می یابد. آنها در گروه های 15-16 تکه ای جمع می شوند و لپه ها را تشکیل می دهند (گروهی از پرزها که به کمک یک تنه پرز به صفحه کوریون متصل می شوند). پرزهای لنگر (در تماس با بافت مادر) و پرزهای انتهایی (در تماس با خون مادر) را اختصاص دهید. بافت همبند تشکیل می شود: سپتوم ها در مرز بین لپه ها.
بخش مادر
BASIC FALLING SHELL (صفحه پایه)
بافت همبند سپتامبر
جای خالی پر شده با خون مادر
سلول های دسیدوال در مخاط بافت همبند ظاهر می شوند (همان سلول های بافت همبند بزرگ و غنی از گلیکوژن هستند. مجرای عروق و هیدراتاسیون مخاط افزایش می یابد، فعالیت فاگوسیتی استثنایی ظاهر می شود.
عملکردهای جفت:
سنتز BJU
یون های کلسیم، آهن، روی، منیزیم، مس، P
رسوب می کند و حاوی ویتامین های گروه های A، B1، B2، B6، C، D، E است
عوامل لخته شدن
تنفسی
تروفیک
غدد درون ریزتمام هورمون ها توسط سمپلاستوتروفوبلاست سنتز می شوند. آنها برای رشد و نمو جنین ضروری هستند، بدن مادر را برای زایمان و شیردهی آماده می کنند.
هورمون ها:
گنادوتروپین جفتی (حفظ جسم زرد ترشح شده از روزهای اول بارداری بر روی جسم زرد اثر می کند و ترشح پروژسترون و استروژن را تحریک می کند که بر روی آندومتر اثر کرده و باعث رشد جنین می شود.
سوماتوماموتروپین کوریونی (بر روی غده پستانی و جسم زرد اثر می کند و باعث تغییراتی در بدن مادر می شود)
کورتیکولیبرین (تاریخ تحویل را مشخص می کند)
حفاظت ایمونولوژیک
مکانیسم های محافظت از جنین: سمپلاستوتروفوبلاست- پروتئین هایی را سنتز می کند که پاسخ ایمنی مادر را مهار می کند. هورمون های جفتی- مهار لنفوسیت های مادر، فیبرینوئید جفتی
مانع (محافظت کننده)- بیان ضعیفی دارد، بنابراین داروها، الکل، نیکوتین، سموم، داروها (آنتی بیوتیک ها، سولفونامیدها، مسکن ها)، ویروس ها، باکتری ها نفوذ می کنند.
سد جفت:سد بین خون مادر و جنین مناسب برای سد: سیتوتروفوبلاست با غشای پایه، بافت همبند، ویلوس استروما، اندوتلیوم عروق خونی و غشای پایه آن. از طرف مادری، خون از طریق 30 شریان مارپیچی رحم با فشار 70-80 میلی متر جیوه وارد فضای بین پرزها می شود. هنر خون مادر پرزها را میشوید و سپس در قسمت انتهایی قسمتهای جفت جمع میشود و از آنجا از طریق سینوس حاشیهای به داخل سیاهرگهای رحم منتقل میشود. حجم خون 150 میلی لیتر.
جنین شناسی شماره 8
مفهوم و معنای جفت. جفت در انسان: ویژگی های زمانی و مورفولوژیکی. نوع و ساختار جفت تشکیل شده.







