Початкові стадії ембріогенезу. Стадії ембріонального розвитку. Чоловічі статеві клітини - сперматозоїди
Ембріогенез (грец. embryon – зародок, genesis – розвиток) – ранній період індивідуального розвитку організму від моменту запліднення (зачаття) до народження, є початковим етапом онтогенезу (грец. ontos – істота, genesis – розвиток), процесу індивідуального розвитку організму від зачаття до смерті.
Розвиток будь-якого організму починається внаслідок злиття двох статевих клітин (гамет), чоловічої та жіночої. Всі клітини тіла, незважаючи на відмінності в будові та виконуваних функціях, поєднує одне – єдина генетична інформація, що зберігається в ядрі кожної клітини, єдиний подвійний набір хромосом (крім вузькоспеціалізованих клітин крові – еритроцитів, які не мають ядра). Тобто всі соматичні (сома - тіло) клітини диплоїдні і містять подвійний набір хромосом - 2 n, і лише статеві клітини (гамети), що формуються в спеціалізованих статевих залозах (насінниках і яєчниках), містять одинарний набір хромосом - 1 n.
При злитті статевих клітин утворюється клітина – зигота, у якій відновлюється подвійний набір хромосом. Нагадаємо, що в ядрі клітини людини міститься 46 хромосом, відповідно статеві клітини мають 23 хромосоми.
Зигота, що утворилася, починає ділитися. І етап поділу зиготи називається дробленням, у результаті якого утворюється багатоклітинна структура морула (тутова ягода). Цитоплазма розподіляється між клітинами нерівномірно, клітини нижньої половини морули більші, ніж верхньої. За обсягом морула можна порівняти з обсягом зиготи.
На II етапі поділу, в результаті перерозподілу клітин, утворюється одношаровий зародок - бластула, що складається з одного шару клітин та порожнини (бластоціль). Клітини бластули різняться між собою за розмірами.
На III етапі, клітини нижнього полюса хіба що вплутуються (инвагинируют) всередину, і утворюється двошаровий зародок - гаструла, що складається із зовнішнього шару клітин - ектодерми та внутрішнього шару клітин - ентодерми.
Дуже скоро, між І і ІІ шарами клітин формується, в результаті поділу клітин, ще один шар клітин, середній - мезодерма, і зародок стає тришаровим. У цьому завершується стадія гаструли.
З цих трьох шарів клітин (їх називають зародковими шарами) формуються тканини та органи майбутнього організму. З ектодерми розвивається покривна та нервова тканина, з мезодерми – скелет, м'язи, кровоносна система, статеві органи, органи виділення, з ентодерми – органи дихання, харчування, печінка, підшлункова залоза. Багато органів формуються з кількох зародкових шарів.
Ембріогенез включає в себе процеси з моменту запліднення до народження.
Розвиток людського організму починається після запліднення жіночої статевої клітини – яйця (ovium) чоловічого – сперматозоїдом (spermatozoon, spermium).
Детальне вивчення розвитку людського зародка (ембріона) є предметом ембріології. Тут ми обмежимося лише загальним оглядом розвитку зародка (ембріогенезу), що необхідне розуміння статури людини.
Ембріогенез всіх хребетних, у тому числі й людини, можна поділити на три періоди.
1. Дроблення: запліднене яйце, spermovium, або зигота послідовно ділиться на клітини (2,4,8,16 і так далі) в результаті чого спочатку утворюється щільна багатоклітинна куля, морула, а потім одношарова бульбашка – бластула, яка містить у середині первинну порожнину, бластоціль. Тривалість цього періоду – 7 днів.
2. Гаструляція полягає у перетворенні одношарового зародка на дво-, а пізніше тришаровий – гаструлу. Перші два шари клітин називаються зародковими листками: зовнішній ектодерма і внутрішній ентодерма (до двох тижнів після запліднення), а третій, середній, що виникає пізніше між ними, шар отримує назву середнього зародкового листка - мезодерми. Другим важливим результатом гаструляції у всіх хордових є виникнення осьового комплексу зачатків: на дорсальному (спинному) боці ентодерми виникає зачаток спинної струни, хорди, а на вентральній (черевній) її стороні – зачаток кишкової ентодерми; на дорзальній стороні зародка, по середній лінії його з ектодерма виділяється нервова пластинка - зачаток нервової системи, а решта ектодерма йде на побудову епідермісу шкіри і тому називається шкірної ектодерма.
Надалі зародок росте у довжину і перетворюється на циліндричну освіту з головним (краніальним) та хвостовим каудальним кінцями. Цей період триває до кінця третього тижня після запліднення.
3. Органогенез і гістогенез: нервова пластинка занурюється під ектодерму і перетворюється на нервову трубку, яка складається з окремих сегментів – невротомів, – і дає початок розвитку нервової системи. Мезодермальні зачатки відшнуровуються від ентодерми первинної кишки і утворюють парний ряд метамерно розміщених мішків, які, розростаючись з боків від тіла зародка, діляться кожен на два відділи: спинний, що лежить з боків від хорди та нервової трубки, і черевної, що лежить з боків від кишки. Спинні відділи мезодерми утворюють первинні сегменти тіла - соміти, кожен з яких у свою чергу ділиться на склерот, який дає початок скелету і міотом, з якого розвивається мускулатура. Із соміту (на бічній його стороні) виділяється також шкірний сегмент – дерматом. Черевні відділи мезодерми, які називаються спланхнотомами, утворюють парні мішки, що містять вторинну порожнину тіла.
Кишкова ентодерма, що залишилася після відокремлення хорди та мезодерми, утворює вторинну кишку – основу для розвитку внутрішніх органів. Надалі закладаються всі органи тіла, матеріалом для побудови яких служать три зародкові листки.
1. Зі зовнішнього зародкового листка, ектодерми, розвиваються:
а) епідерміс шкіри та її похідні (волосся, нігті, шкірні залози);
б) епітелій слизової оболонки носа, рота та заднього проходу;
в) нервова система та епітелій органів чуття.
2. З внутрішнього зародкового листка, ентодерми, розвивається епітелій слизової оболонки більшої частини травного тракту з усіма залізистими структурами, що належать сюди, більшій частині дихальних органів, а також епітелій щитовидної та зобної залоз.
3. З середнього зародкового листка, мезодерми, розвивається мускулатура скелета, мезотелій оболонок серозних порожнин із зачатками статевих залоз та нирок.
Крім того, зі спинних сегментів мезодерми виникає ембріональна сполучна тканина, мезенхіма, яка дає всі види сполучної тканини, у тому числі хрящову та кісткову. Оскільки спочатку мезенхіма проводить поживні речовини до різних ділянок зародка, виконуючи трофічну функцію, пізніше з неї розвиваються кров, лімфа, кровоносні судини, лімфатичні вузли, селезінка.
Крім розвитку самого зародка, необхідно враховувати також утворення позазародкових частин, за допомогою яких ембріон отримує необхідні для життя поживні речовини.
У багатоклітинній щільній кулі виділяється внутрішній зародковий вузлик, ембріобласт, і зовнішній шар клітин, який відіграє важливу роль у харчуванні зародка і тому називається трофобласт. За допомогою трофобласта зародок проникає в товщу слизової оболонки матки (вживлення), і тут починається утворення особливого органу, за допомогою якого встановлюється зв'язок зародка з тілом матері та здійснюється його харчування. Цей орган називається дитячим місцем, послідом, або плацентою. Ссавці, які мають плаценту називаються плацентарними. Поруч із утворенням плаценти йде процес відокремлення зародка, що розвивається, від позазародкових частин у результаті виникнення так званої тулубової складки, яка, вдаючись гребенем до середини, ніби відшнуровує кільцем тіло зародка від позазародкових частин. При цьому, однак, зберігається з'єднання з плацентою за допомогою стебла пупкового, який далі перетворюється на пупковий канатик. На ранніх стадіях розвитку в останньому проходить жовткова протока, яка з'єднує кишку з її випинанням у позазародкову ділянку – жовтковий мішок. У хребетних, які не мають плаценти, жовтковий мішок містить поживний матеріал яйця – жовток і є важливим органом, через який здійснюється харчування зародка.
У людини жовтковий мішок хоч і виникає, але помітну роль розвитку зародка не грає і після всмоктування його вмісту поступово редукується. У пупковому канатиці проходять пупкові (плацентарні) судини, через які тече кров від плаценти в тіло зародка і назад. Вони розвиваються з мезодерми сечового мішка, або алантоіса, який випирається з вентральної стінки кишки і виходить із тіла зародка через пупковий отвір у позазародкову частину. У людини з частини алантоіса, що міститься в середині тіла зародка, утворюється частина сечового міхура, та якщо з його судин утворюються пупкові кровоносні судини. Зародок, що розвивається, покритий двома зародковими оболонками. Внутрішня оболонка, амніон, утворює об'ємний мішок, який наповнений білковою рідиною і утворює рідке середовище для зародка, через що мішок називають водною оболонкою. Весь зародок разом з амніотичним та жовтковим мішками оточений зовнішньою оболонкою (до складу якої входить і трофобласт). Ця оболонка, маючи ворсинки, називається ворсинчастою, або хоріон. Хоріон виконує трофічну, дихальну, видільну та бар'єрну функції.
13. На відміну від мохів, у папоротей, хвощів і плаунів у циклі розвитку переважає спорофіт - листостебельна рослина. Представники цих трьох груп рослин мають листя, стебла та коріння. У більшості з них є підземні кореневища з видозміненим листям і підрядним корінням. Сучасні хвощі, плауни та папороті - в основному трав'янисті рослини. Тільки в тропіках і субтропіках тут деревоподібні папороті. Однак у стародавні епохи - 200-350 млн років тому ці групи рослин були представлені деревоподібними формами і становили дрімучі ліси, що дали початок найбільшим кам'яним.
вугільних відкладень світу (Донбас, Кузбас та ін.).
Які особливості будови плаунів, хвощів та папоротей?
Розглянемо особливості плаунів, хвощів та папоротей. Сучасні плауноподібні - багаторічні, зазвичай вічнозелені трави. Найвідоміший представник плауноподібних - плаун звичайний, поширений у середній смузі Росії в сирих ялинових і
соснових лісів. Це рослина з гнучким розгалуженим стеблом, що стелиться по землі. Листя дрібне, розташоване на стеблі по спіралі. Наприкінці літа на бічних гілочках з'являються зазвичай два спороносні колоски. Кожен колосок утворений дрібним тонким видозміненим листям, яке називаються спорофілами. В основі спорофілів розташовані спорангії, де формуються спори. Хвощі, або хвощевидні, легко відрізнити за членистою будовою стебел: вони яскраво виражено чергування вузлів і междоузлий. Листя на стеблі розташовуються мутовками (по кілька штук у вузлі), оточуючи стебло. На верхівках стебел формуються спороносні колоски, в яких дозрівають суперечки. У деяких видів, наприклад у хвоща польового, стебла бувають двох типів: спороносні (буро рожеві, розвиваються навесні і після спороношення відмирають) і вегетативні (з'являються влітку від того ж самого кореневища). Папоротеподібні представлені в природі багаторічними травами, ліанами, деревами та епіфітами, що поселяються на стовбурах дерев. У папороті велике листя; молоді зазвичай згорнуті у вигляді равликів. Папороті нашої країни мають кореневища. Спорангії у них розташовуються на нижній стороні листа і зібрані в купки - їх називають сорусами. У вимерлих папоротей спорангії були одиночними. На території нашої країни ростуть папороті орляк звичайний, щитовник чоловічий, кочедижник жіночий, багатоніжка звичайна та інші види.
У чому особливість розвитку папоротей, хвощів та плаунів?
Розмноження у всіх трьох груп вищих спорових рослин відбувається за однією схемою. Розглянемо його на прикладі папороті. На нижній стороні листка дорослої рослини розвиваються спорангії зі суперечками. Потрапляючи у сприятливі умови, суперечка проростає та дає початок гаметофіту. Він має вигляд маленької платівки з ризоидами і називається заростком. На заростку розвиваються чоловічі та жіночі гаметангії з
статевими клітинами - яйцеклітинами та сперматозоїдами. Після запліднення, яке відбувається за наявності води, із зиготи спочатку розвивається зародок, а потім і доросла рослина - спорофіт. Таким чином, у плаунів, хвощів і папоротей має місце чергування статевого покоління (заросток - гаметофіт) і безстатевого (зростав).
ла рослина - спорофіт).
14. Післязародковий розвиток: прямий і непрямий. Причини ослаблення конкуренції між батьками та потомством при непрямому розвитку.
1. Індивідуальний розвиток організму (онтогенез) - період життя, який при статевому розмноженні починається з утворення зиготи, характеризується незворотними змінами (збільшенням маси, розмірів, появою нових тканин та органів) та завершується смертю.
2. Зародковий (ембріональний) та післязаро-диховий (постембріональний) періоди індивідуального розвитку організму.
3. Післязародковий розвиток (приходить на зміну зародковому) - період від народження або виходу зародка з яйця до смерті. Різні шляхи післязародкового розвитку тварин - пряме та непряме:
1) Прямий розвиток - народження потомства, зовні схожого на дорослий організм. Приклади: розвиток риб, плазунів, птахів, ссавців, деяких видів комах. Так, малеча риби схожа на дорослу рибу, каченя на качку, кошеня на кішку;
2) непрямий розвиток - народження або вихід з яйця потомства, що відрізняється від дорослого організму за морфологічними ознаками, спосіб життя (типу харчування, характер пересування). Приклад: з яєць хруща з'являються червоподібні личинки, живуть у грунті і живляться корінням на відміну від дорослого жука (живе на дереві, харчується листям).
Стадії непрямого розвитку комах: яйце, личинка, лялечка, доросла особина. Особливості життя тварин на стадії яйця та лялечки – вони нерухомі. Активний спосіб життя личинки та дорослого організму, різні умови проживання, використання різної їжі.
4. Значення непрямого розвитку - ослаблення конкуренції між батьками і потомством, оскільки вони поїдають різну їжу, вони різні місця проживання. Непрямий розвиток - важливий пристрій, що виник у процесі еволюції. Воно сприяє ослабленню боротьби за існування між батьками та потомством, виживання тварин на ранніх стадіях післязародкового розвитку.
Загальна характеристика. Перші голонасінні з'явилися наприкінці девонського періоду близько 350 млн років тому; ймовірно, вони походять від древніх папоротеподібних, які вимерли на початку кам'яновугільного періоду. У мезозойську епоху - епоху гороутворення, підняття материків і висушення клімату - голонасінні досягли розквіту, але з середини крейдяного періоду поступилися своє панівне становище покритонасінним.
Відділ сучасних голонасінних налічує понад 700 видів. Незважаючи на відносно малу чисельність видів, голонасінні завоювали майже всю земну кулю. У помірних широтах Північної півкулі вони у величезних просторах утворюють хвойні ліси, звані тайгою.
Сучасні голонасінні представлені переважно деревами, значно рідше – чагарниками та дуже рідко – ліанами; трав'янистих рослин у тому числі немає. Листя голонасінних значно відрізняється від інших груп рослин не тільки за формою та розмірами, але й за морфологією та анатомією. У більшості видів вони голкоподібні (хвоя) або лускоподібні; у окремих представників вони великі (наприклад, у вельвічії дивовижна їх довжина досягає 2-3 м), перисторозсічені, дволопатеві та ін. Листя розташовуються поодинці, по два або кілька в пучках.
Переважна більшість голонасінних - вічнозелені, одно-або дводомні рослини з добре розвиненими стеблами та кореневою системою, утвореною головним і бічним корінням. Розселяються вони насінням, яке формується із сім'язачатків. Сім'язачатки голі (звідси назва відділу), розташовані на мегаспорофілах або насіннєвих лусках, зібраних в жіночі шишки.
У циклі розвитку голонасінних спостерігається послідовна зміна двох поколінь - спорофіту та гаметофіту з пануванням спорофіту. Гаметофіти сильно редуковані, причому чоловічі гаметофіти голо- і покритонасінних рослин не мають антеридіїв, чим різко відрізняються від усіх різноспорових безнасінних рослин.
Голосонасінні включають шість класів, два з яких повністю зникли, а інші представлені рослинами, що нині живуть. Найбільш збереженою і найчисленнішою групою голонасінних є клас Хвойні, що налічує не менше 560 видів, що утворюють ліси на великих просторах Північної Євразії та Північної Америки. Найбільше видів сосни, ялини, модрини зустрічається біля побережжя Тихого океану.
Клас Хвойні. Всі хвойні - вічнозелені, рідше листопадні (наприклад, модрина) дерева або чагарники з голчастим або лускоподібним (наприклад, у кипарису) листям. Голкове листя (хвоя) щільне, шкірясте і жорстке, покрите товстим шаром кутикули. Продихи занурені в поглиблення, заповнені воском. Всі ці особливості будови листя забезпечують хороше пристосування хвойних до зростання як у посушливих, так і в холодних місцеперебуваннях.
У хвойних прямостоячі стовбури, вкриті лускатою корою. На поперечному розрізі стебла добре видно розвинена деревина та менш розвинені кора та серцевина. Ксилема хвойних на 90-95% утворена трахеїдами. Шишки хвойних роздільностатеві; рослини – частіше однодомні, рідше – дводомні.
Найбільш широко поширеними представниками хвойних у Білорусі та Росії є сосна звичайна та ялина звичайна, або європейська. Їхня будова, розмноження, чергування поколінь у циклі розвитку відображає характерні особливості всіх хвойних.
Сосна звичайна – однодомна рослина (рис. 9.3). У травні біля основи молодих пагонів сосни утворюються пучки зеленувато-жовтих чоловічих шишок довжиною 4-6 мм і діаметром 3-4 мм. На осі такої шишки розташовані багатошарові лускаті листочки, або мікроспорофіли. На нижній поверхні мікроспорофілів знаходяться два мікроспорангії - пилкових мішка, в яких утворюється пилок. Кожне пилкове зерно забезпечене двома повітряними мішками, що полегшує перенесення пилку вітром. У пилковому зерні є дві клітини, одна з яких згодом, при попаданні на сім'язачаток, формує пилкову трубку, інша після поділу утворює два спермію.
На інших пагонах тієї ж рослини утворюються жіночі шишки червоного кольору. На їхній головній осі розташовуються дрібні прозорі лусочки, що криють, у пазухах яких сидять великі товсті, згодом здерев'янювальні луски. На верхній стороні цих лусок розташовано по два сім'язачатки, у кожному з яких розвивається жіночий гаметофіт - ендосперм із двома архегоніями з великою яйцеклітиною у кожному з них. На верхівці сім'язачатка, зовні захищеного інтегументом, є отвір - пилковхід, або мікропиле.
Пізньої весни або на початку літа дозрілий пилок розноситься вітром і потрапляє на сім'ячаток. Через мікропилку пилок втягується всередину сім'язачатка, де і проростає в пилкову трубку, яка проникає до архегоній. Два спермію по пилковій трубці, що утворилися до цього часу, потрапляють до архегоній. Потім один із сперміїв зливається з яйцеклітиною, а інший відмирає. З заплідненої яйцеклітини (зиготи) формується зародок насіння, а сім'язачаток перетворюється на насіння. Насіння у сосни дозрівають на другий рік, висипаються з шишок і, підхоплені тваринами або вітром, переносяться на значні відстані.
За своїм значенням у біосфері та ролі у господарській діяльності людини хвойні займають друге місце після покритонасінних, далеко перевершуючи всі інші групи вищих рослин.
Вони допомагають вирішувати величезні водоохоронні та ландшафтні завдання, служать найважливішим джерелом деревини, сировини для одержання каніфолі, скипидару, спирту, бальзамів, ефірних олій для парфумерної промисловості, лікарських та інших цінних речовин. Деякі хвойні культивуються як декоративні (ялиці, туї, кипариси, кедри та ін.). Насіння ряду сосен (сибірської, корейської, італійської) вживається в їжу, з них також отримують олію.
Представники інших класів голонасінних (саговникові, гнетові, гінкгові) зустрічаються значно рідше і менш відомі, ніж хвойні. Однак майже всі види саговникових декоративні та користуються широкою популярністю у садівників багатьох країн. Вічнозелені безлисті невисокі чагарники ефедри (клас гнетових) є джерелом сировини для отримання алкалоїду ефедрину, який застосовується як засіб, що збуджує центральну нервову систему, а також при лікуванні захворювань алергічного характеру.
16. Радіальна симетрія- Форма симетрії, при якій тіло (або фігура) збігається саме з собою при обертанні об'єкта навколо певної точки або прямої. Часто ця точка збігається з центром симетрії об'єкта, тобто тією точкою, де перетинається нескінченна кількість осей двосторонньої симетрії. Радіальну симетрію мають такі геометричні об'єкти, як коло, куля, циліндр або конус.
У біології про радіальну симетрію говорять, коли через тривимірну істоту проходять одна чи більше осей симетрії. При цьому радіальносиметричні тварини можуть і не мати площин симетрії. Так, у сифонофори Velella є вісь симетрії другого порядку і немає площин симетрії
Зазвичай через вісь симетрії проходять дві чи більше площини симетрії. Ці площини перетинаються прямою - осі симетрії. Якщо тварина обертатиметься навколо цієї осі на певний градус, то вона буде відображатися сама на собі (збігатися сама з собою).
Таких осей симетрії може бути кілька (поліаксонна симетрія) або одна (монаксонна симетрія). Поліаксонна симетрія поширена серед протистів (наприклад, радіолярій).
Як правило, у багатоклітинних тварин два кінці (полюси) єдиної осі симетрії нерівноцінні (наприклад, у медуз на одному полюсі (оральному) знаходиться рот, а на протилежному (аборальному) – верхівка дзвону. Така симетрія (варіант радіальної симетрії) у порівняльній анатомії називається одновісно-гетеропольної У двовимірній проекції радіальна симетрія може зберігатися, якщо вісь симетрії спрямована перпендикулярно до проекційної площини, тобто збереження радіальної симетрії залежить від кута спостереження.
Радіальна симетрія характерна для багатьох куховарців, а також для більшості голкошкірих. Серед них зустрічається так звана пентасиметрія, що базується на п'яти площинах симетрії. У голкошкірих радіальна симетрія вторинна: їх личинки двосторонньосиметричні, а у дорослих тварин зовнішня радіальна симетрія порушується наявністю мадрепорової платівки.
Крім типової радіальної симетрії існує двопроменева радіальна симетрія (дві площини симетрії, наприклад, у гребенів). Якщо площина симетрії лише одна, то симетрія билатеральная (таку симетрію мають тварини групи Bilateria).
У квіткових рослин часто зустрічаються радіальносиметричні квітки: 3 площини симетрії (водокрас жаб'ячий), 4 площини симетрії (перстач прямий), 5 площин симетрії (дзвіночок), 6 площин симетрії (безвременник). Квітки з радіальною симетрією називаються актиноморфні, квітки з білатеральною симетрією – зигоморфні.
Білатеральна симметрія(двостороння симетрія) - симетрія дзеркального відображення, при якій об'єкт має одну площину симетрії, щодо якої дві половини його дзеркально симетричні. Якщо на площину симетрії опустити перпендикуляр з точки A і потім з точки О на площині симетрії продовжити його на довжину AО, то він потрапить у точку A1, у всьому подібну до точки A. Вісь симетрії у білатерально симетричних об'єктів відсутня. У тварин білатеральна симетрія проявляється у схожості або майже повній ідентичності лівої та правої половин тіла. При цьому завжди існують випадкові відхилення від симетрії (наприклад, відмінності в папілярних лініях, розгалуженні судин та розташування родимок на правій та лівій руках людини). Часто існують невеликі, але закономірні відмінності у зовнішній будові (наприклад, більш розвинена мускулатура правої руки у праворуких людей) та більш суттєві відмінності між правою та лівою половиною тіла у розташуванні внутрішніх органів. Наприклад, серце у ссавців зазвичай розміщене несиметрично, зі зміщенням вліво.
У тварин поява білатеральної симетрії в еволюції пов'язана з повзанням по субстрату (дном водойми), у зв'язку з чим з'являються спинна і черевна, а також права і ліва половини тіла. Загалом серед тварин білатеральна симетрія більш виражена у активно рухливих форм, ніж у сидячих. Білатеральна симетрія властива всім досить високоорганізованим тваринам, крім голкошкірих. В інших царствах живих організмів білатеральна симетрія властива меншій кількості форм. Серед протистів вона характерна для дипломонад (наприклад, лямблій), деяких форм трипаносом, бодонідом, раковинками багатьох форамініферів. У рослин білатеральну симетрію має зазвичай не весь організм, яке окремі частини - листя чи квітки. Білатерально-симетричні квітки ботаніки називають зигоморфними.
17. Покритонасінні (квіткові, маточкові)за часом появи Землі є наймолодшою і водночас найбільш високоорганізованою групою рослин. У процесі еволюції представники цього відділу з'явилися пізніше за інших, але вони дуже швидко зайняли панівне становище на земній кулі.
Найбільш характерною відмінністю покритонасінних є наявність у них своєрідного органу - квітки, який відсутній у представників інших відділів рослин. Тому покритонасінні і називаються частіше ще квітковими рослинами. Семяпочка у них прихована, вона розвивається всередині маточка, в його зав'язі, тому покритонасінні називаються інакше маточними. Пилок у покритонасінних вловлюється не сім'япочками, як у голонасінних, а особливою освітою - приймочкою, яким закінчується маточка.
Після запліднення яйцеклітини з семяпочки утворюється насіння, а зав'язь розростається в плід. Отже, насіння у покритонасінних розвивається в плодах, тому цей відділ рослин і називається покритонасінним.
Покритонасінні (Angiospermae), або квіткові (Magnoliophyta) – відділ найбільш досконалих вищих рослин, що мають квітку. Раніше включалися до відділу насіннєвих рослин разом із голонасінними. На відміну від останніх сім'язачатки квіткових укладені в зав'язь, утворену плодолистками, що зрослися.
Квітка є генеративним органом покритонасінних рослин. Він складається з квітконіжки та квітколожа. На останньому розташовуються оцвітина (простий або подвійний), андроцей (сукупність тичинок) та гінецей (сукупність плодолистків). Кожна тичинка складається з тонкої нитки тичинки і розширеного пильовика, в якому дозрівають спермії. Плодолист квіткових рослин представлений маточкою, що складається з масивної зав'язі та довгого стовпчика, вершинна розширена частина якого називається рильце.
Покритонасінні мають вегетативні органи, що забезпечують механічну опору, транспорт, фотосинтез, газообмін, а також запасання поживних речовин, і генеративні органи, що беруть участь у статевому розмноженні. Внутрішня будова тканин найскладніше з усіх рослин; ситоподібні елементи флоеми оточені клітинами-супутницями; Багато представників покритонасінних мають судини ксілеми.
Чоловічі гамети, що містяться всередині пилкових зерен, потрапляють на рильце і проростають. Гаметофіти квіткових вкрай спрощені та мініатюрні, що значно скорочує тривалість циклу розмноження. Утворюються вони в результаті мінімальної кількості мітозів (трьох у жіночого гаметофіту та двох у чоловічого). Одна з особливостей статевого розмноження – подвійне запліднення, коли один із сперміїв зливається з яйцеклітиною, утворюючи зиготу, а другий – з полярними ядрами, утворюючи ендосперм, який є запасом поживних речовин. Насіння квіткових рослин укладено в плід (звідси їх друга назва - покритонасінні).
18. ДНКЯвище це було відкрито у дослідах із пневмококами, тобто з бактеріями, що викликають запалення легень. Відомі дві форми пневмококів: А-форма з полісахаридною капсулою та Б-форма без капсули. Обидві ці ознаки спадкові.
Пневмококи А-форми при зараженні ними мишей викликають запалення легень, від якого миші гинуть. Б-форма їм нешкідлива.
У 1928 році англійський бактеріолог Ф. Гріффітс заражав мишей сумішшю, що складається з вбитих нагріванням пневмококів А-форми та живих пневмококів Б-форми. Вчений припускав, що миші не захворіють. Але всупереч очікуванням піддослідні тварини загинули. Ф. Гріффітсу вдалося виділити з тканин загиблих мишей пневмококи. Усі вони виявилися капсульованими, тобто А-форми. Отже, убита форма якимось чином передавала свої властивості живим клітин Б-форми. Але як? За допомогою якої саме речовини: полісахариду, з якої складається капсула, білка чи ДНК?
Від вирішення цього питання залежало багато, оскільки, встановивши речовину, що передає спадкову ознаку - утворення капсули, можна було отримати відповідь. Однак зробити це не вдавалося досить довго. Лише через 16 років після дослідів Ф. Гріффітса, в 1944 році, американський вчений А. Евері зі співробітниками, поставивши ряд чітких експериментів, зумів з повним обґрунтуванням довести, що полісахарид та білок не мають жодного відношення до передачі спадкових властивостей пневмокока А-форми.
В процесі цих експериментів за допомогою спеціального ферменту розчинили полісахаридну капсулу вбитих пневмококів А-форми і перевірили, чи залишки клітини форми А передавати спадкову інформацію клітинам форми Б. Виявилося, що продовжують. Стало ясно, що полісахарид як джерело генетичної інформації відпадає.
Таким чином, методом виключення було встановлено, що спадкову інформацію в клітині зберігає та передає молекула ДНК. І справді, коли зруйнували ДНК, утворення капсульних форм А з безкапсульних Б припинилося.
Явище перетворення, тобто спадкової зміни властивостей однієї форми бактерій під впливом речовин іншої форми, було названо трансформацією. Речовина, що викликає трансформацію, отримала назву трансформуючого агента. Їм, як було встановлено, є ДНК.
Кожен білок представлений одним або декількома поліпептидними ланцюгами. Ділянка ДНК, що несе інформацію про один поліпептидний ланцюг, називається геном. Кожна молекула ДНК містить багато різних генів. Сукупність молекул ДНК клітини виконує функцію носія генетичної інформації. Завдяки унікальній властивості - здатності до подвоєння, яким не має жодна інша з відомих молекул, ДНК можуть копіюватися. При розподілі "копії" ДНК розходяться по двох дочірніх клітинах, кожна з яких внаслідок цього матиме ту саму інформацію, яка містилася в материнській клітині. Оскільки гени - це ділянки молекул ДНК, дві клітини, що утворюються при розподілі, мають однакові набори генів. Кожна клітина багатоклітинного організму при статевому розмноженні виникає з однієї заплідненої яйцеклітини внаслідок багаторазових поділів. Отже, помилка, що випадково виникла в гені однієї клітини, буде відтворена в генах мільйонів її нащадків. Ось чому всі еритроцити хворого на серповидноклітинну анемію мають однаково зіпсований гемоглобін. Помилка сталася в гені, що несе інформацію про бета-ланцюг білка. Копією гена є і-РНК. Нею, як у матриці, у кожному еритроциті тисячі разів " друкується " неправильний білок. Діти отримують зіпсовані гени від батьків через їхні статеві клітини. Генетична інформація передається як від однієї клітини до дочірніх клітин, так і від батьків дітям. Ген є одиницею генетичної, чи спадкової інформації.
Ембріологія(Від грец. Embryon - зародок і logos - слово, наука) - наука про розвиток зародка. Вона вивчає період індивідуального розвитку, який починається заплідненням та закінчується (у ссавців та людини) народженням плода.
Ембріологія вивчає морфологічні прояви освіти із заплідненої яйцеклітини (зиготи) складного багатоклітинного організму, до складу якого входять чотири тканини, кілька десятків клітинних диферонів, органи, системи органів та апарати та частини тіла.
Знання з ембріології сприяють кращому засвоєнню макроскопічної анатомії. Вибудовуючись у логічний ряд з ембріологічними даними, анатомічні факти, що стосуються макро- та мікроструктури організму, підтверджують, що структурні залежності в постнатальному періоді розвитку – закономірний результат процесів морфогенезу, що відбуваються у зародках.
Ембріологія дозволяє визначити морфологічні прояви, повноцінність реалізації генетичної програми, закладеної в зиготі, а також порушення цієї реалізації, що проявляється у структурних та функціональних аномаліях ембріонального розвитку, які нерідко зустрічаються у клінічній практиці та пояснити які можна, лише знаючи ембріологію.
Процес ембріонального розвитку людини, як та інших хребетних, ділиться на ряд стадій, що характеризуються якісними та кількісними особливостями. Головними стадіями ембріогенезу є запліднення, дроблення, гаструляція, утворення тканин (гістогенез), органів (органогенез), а також систем органів та апаратів (системогенезу). У деяких підручниках та посібниках до ембріогенезу відносять також прогенез – розвиток жіночих та чоловічих статевих клітин.
У цьому розділі розглянуто, головним чином, початкові етапи розвитку зародка людини, що починається заплідненням та завершується утворенням зародків тканин та позазародкових органів. Що стосується гісто-, органо- та системогенезу, то вони будуть розглядатися при вивченні будови відповідних органів та систем.
Запліднення є злиттям чоловічої та жіночої статевих клітин, що призводить до утворення одноклітинного зародка - зиготи, що містить диплоїдний набір хромосом і поєднує материнську та батьківську спадковість. Запліднення відбувається у яйцеводах. Тривалість процесу – близько доби.
Зигота починає ділитися шляхом мітозу. Її поділ називається дробленням, бо в процесі послідовного поділу зиготи клітини зменшуються. У зв'язку з цим загальні розміри багатоклітинного зародка спочатку можуть бути приблизно такими ж, як і зиготи.
Клітини, що утворюються в процесі дроблення, отримали назву бластомерів (від грец. Blastos – зачаток, meros – частина). Період дроблення починається з поділу зиготи на два бластомери і продовжується аж до утворення бульбашки (бластули). У людини цей період починається через добу після запліднення та триває приблизно 4-5 діб.
Одна з особливостей дроблення зиготи ссавців та людини – виникнення бластомерів, які різняться між собою величиною та швидкістю дроблення. Зовні розташовані дрібні, світлі бластомери (мікроміри), що скоріше діляться і утворюють у сукупності трофобласт, або живий зачаток (від грец. Tropho – живлю). У центрі зародка містяться великі темні бластомери (макроміри) з базофільною цитоплазмою, діляться повільніше і утворюють в сукупності ембріобласт, або зародок зародка.
Зародок, не має порожнини, називається морули (від грец. Morula – ягода шовковиці). Хоча цей термін характерний для ранніх зародків хордових (наприклад, ланцетника, який насправді нагадує на початкових етапах розвитку ягоду шовковиці), він вживається також для позначення ранніх зародків вищих хребетних, які ще не мають порожнини і є щільним скупченням клітин.
Потім у зародку з'являється невелика, заповнена рідиною порожнину і він набуває вигляду кульки, яка називається бластоцистою (бластули). Стінка бластоцисти утворена трофобластом. На внутрішній поверхні зародка розташований ембріобласт.
У середині шостого дня у зародку налічується понад сто клітин. У цей період він знаходиться в матці, де відбувається його імплантація (від лат. Inrlаntatio - проникнення), тобто заглиблення в слизову оболонку. Кількість бластомерів у зародку у своїй збільшується.
Наприкінці першого тижня розвитку з трофобласту у процесі імплантації утворюється двошаровий епітелій, внутрішній шар якого називається цитотрофобластом. Він представлений клітинами кубічної форми. Зовні знаходиться плазмодиотрофобласт, який називають також синцитіотрофобласти, він є симпластами, тобто. неподілену на клітинні території цитоплазматичну масу із численними ядрами.
Проникнення зародка в слизову оболонку матки пов'язане з дією трофобласту, що виділяє гістолітичні ферменти, що зумовлюють локальне (місцеве) руйнування внутрішньої оболонки матки – ендометрію. Зародок поринає у його товщу і ендометрій стуляє з нього. Таким чином, подальший розвиток ембріона відбувається у слизовій оболонці матки.
Трофобласт забезпечує харчування зародка на ранніх етапах його розвитку, бере активну участь в імплантації зародка і у формуванні тимчасового органу - хоріона який, взаємодіючи зі слизовою оболонкою матки, утворює плаценту - позазародковий орган, який забезпечує зв'язок зародка з організмом матері. Розвиток, будова та функціональне значення плаценти розглядаються в курсах ембріології та гістології.
З клітинних елементів ембріобласта розвивається зародок та інші позазародкові органи. Одна з суттєвих особливостей розвитку людини – дуже раннє утворення позазародкової мезодерми. Джерело її виникнення – ембріобласт. З нього виселяються клітини, як смужок заповнюють порожнину бластоцист і вистилають зсередини трофобласт. Позародкова мезодерма швидко перетворюється на позазародкову мезенхіму, внаслідок чого зовнішня оболонка зародка складається з трофобластичного епітелію та мезенхіми. Вона називається хоріоном. Як зазначалося вище, хоріон утворює зародкову частину плаценти - органу, що забезпечує зв'язок зародка з організмом матері.
Потім починається гаструляція, яка поділяється на два етапи. У першому утворюються зародкові листки, але в другому - осьовий комплекс, визначальний загальний план будови тіла хребетних і людини і зародкових зачатків, у тому числі утворюються різні тканини.
Зародковими листками називають первинні пласти клітин, що утворюються у першій фазі гаструляції. Вони відмінні за розташуванням (зовнішній листок – ектодерма, внутрішній – ентодерма та середній – мезодерма), розмірами та будовою клітин та спрямованістю подальшого розвитку. До осьового комплексу відносяться нервова трубка, хорда (спинна струна), дорсальна мезодерма, розташована з боків від нервової трубки та хорди, первинна кишка. У другій фазі гаструляції утворюються також ембріональні зачатки - комплекси клітин, що виникають із зародкових листків та починають різні тканини. Таким чином, з того самого ембріонального листка утворюються різні тканини.
Морфологічним проявом першої фази гаструляції, що спостерігається у 7,5-тижневого зародка, є поділ ембріобласту на два шари – зовнішній (епібласт) та внутрішній (гіпобласт).
Епібласт має зачатки ектодерми, зародкової мезодерми, хорди (спинної струни) та зародкової (кишкової) ентодерми. Гіпобласт є зачатком позазародкової або жовткової ентодерми, що є частиною внутрішнього зародкового листка, яка згодом увійде до складу одного з тимчасових органів - жовткового мішка.
Ендодермальні клітини, що відокремилися від епібласту, починають обростати навколо порожнини, що утворилася в розташованих нижче ділянках позазародкової мезодерми. Поступово вони набувають вигляду чаші, краї якої зростаються, внаслідок чого утворюється жовтковий пляшечку. Цей процес з початком утворення жовткового мішка аналогічний тому, що характеризує розвиток цього позазародкового органу у ссавців.
Згодом після початку імплантації в епібласті з'являється первинна амніотична порожнина за рахунок появи рідини, яка розсуває клітини. Потім покрівля цієї порожнини (амніотичного пухирця) розкривається і тимчасово усувається ззовні ділянкою цитотрофобласту. Після цього стінки амніотичної бульбашки ростуть вгору і зливаються, внаслідок чого порожнина амніотичної бульбашки знову стає замкненою, оточеною клітинами епібласту. Цей процес нагадує виникнення так званих амніотичних складок у птахів та ссавців.
З 9 до 14 діб розвитку зародок людини має таку будову: зовнішня стінка, утворена хоріоном, складається з позазародкової мезенхіми та трофобластичного епітелію. Останній, як зазначалося раніше, включає до свого складу цитотрофобласт та плазмодіотрофобласт. У потовщеній стінці зародка, яка звернена вглиб стінки матки, у позазародковій мезенхімі містяться дві бульбашки, які контактують між собою, - амніотичний та жовтковий. Ділянка амніотичного пляшечки, прилеглий до жовткового, утворює потовщення - так званий зародковий щиток, з якого формується тіло зародка. Від мезенхіми, що обмежують дві бульбашки, до хоріону тягнеться смужка клітин - амніотична ніжка, місце відходження якої відповідає задньому кінці тіла зародка.
Завдяки описаним вище процесам зародок забезпечується трьома позазародковими органами – хоріоном, що бере участь у освіті плаценти, амніоном та жовтковим мішком. З більшої частини амніотичного бульбашки формується позазародковий орган - амніон, однією з функцій якого є утворення амніотичної рідини - штучного водного середовища для зародка, що розвивається. З гіпобласта утворюється жовтковий пляшечку (мішок), який не міститься у людини і більшості ссавців жовтка, але виконує важливу роль першого органу кровотворення, що бере участь у розвитку первинних статевих клітин.
Таким чином, у процесі ембріонального розвитку людини поява низки позазародкових органів (хоріону, амніону та жовткового мішка) передує виникненню тіла зародка, що є свідченням першочергового створення умов, необхідних для розвитку ембріона.
На 15 добу внутрішньоутробного розвитку починається друга фаза гаструляції, яка відбувається однаково як у птахів, так і у плацентарних ссавців. Перехід від першої фази гаструляції до другої здійснюється поступово, поєднуючи ознаки першої (утворення зародкових листків) та другої (формування комплексу осьових органів). Зовнішньою ознакою другої фази гаструляції є поява в задньому кінці зародкового щитка витягнутої в довжину смужки клітин, яка називається первинною смужкою. На передньому її кінці утворюється невелике підняття клітин – первинний, чи гензеновський вузлик.
На вершині первинного вузлика з'являється невелика заглибина - первинна ямка. У центральній частині первинної смужки з'являється первинна борозенка, яка є продовженням первинної ямки. Частина матеріалу зовнішнього зародкового листка через первинну ямку занурюється всередину і входить до складу переднього відділу ентодерми, утворюючи так звану передхордальну пластинку, з якої утворюється епітеліально вистелений передній відділ травного каналу та дві передні пари соміту (сегментованих ділянок дорсальної мезодерми).
Орієнтація первинної смужки визначає розташування осьових органів і, таким чином, загальний план будови тіла хребетних.
Частина клітин епібласту мігрує через первинну смужку і, проникнувши в підлеглу частину гіпобласта, утворює зачаток кишкової ентодерми, яка згодом бере участь у розвитку кишки. Друга частина гіпообласту, як зазначалося вище, йде на утворення жовткової ентодерми – внутрішнього шару жовткового мішка.
Внаслідок міграції клітин з епібласту через первинну смужку також виникає зародкова мезодерма, яка розширюється у латеральних напрямках під епібласт.
З утворенням зародкової мезодерми тісно пов'язане походження спинної струни – хорди. Її зачаток утворюється у вигляді так званого хордального (або головного) відростка – тяж клітин, що утворилися за рахунок зародкової мезодерми, що росте вперед від первісного (гензенівського) вузлика між епібластом та гіпобластом. Хорда визначає місцезнаходження майбутнього хребетного стовпа, оскільки навколо неї формуються хребці, та суттєво впливає на розвиток нервової системи. У вищих ссавців залишки хорди зберігаються й у постнатальном періоді розвитку у складі драглистого ядра, nucleus pulposus, міжхребцевих дисків.
Верхній шар клітин зародкового щитка після утворення хорди перетворюється на ектодерму. Під впливом хорди або хордомезодерми на ектодерму, що міститься над нею, остання потовщується і перетворюється на нервову платівку, потім подовжений нервовий клубок, а надалі - в нервову трубку, за рахунок якої утворюється як центральна, так і периферична нервова система.
Наприкінці другого - початку третього тижня ембріонального розвитку закладається четвертий позазародковий орган - алантоїс. Спочатку він має вигляд виросту жовткового мішка, сліпо закінчується та вростає в амніотичну ніжку. Пізніше (після утворення кишки) алантоїс відходить від передньої стінки заднього відділу кишкової трубки. Слідом за утворенням нервової трубки шари мезодерми, що лежать з обох боків від неї і від хорди, підлягають сегментації. Латеральні відділи мезодерми, які пізніше опиняються у складі черевної частини зародка і названі черевною мезодермою, розщеплюються на два шари: зовнішній, контактуючий з ектодермою (Парієтальний листок) і внутрішній, з'єднаний з ентодермою (вісцеральний лист. Сомити з'єднані з вентральною мезодермою тонкими тяжами клітин, званих проміжною мезодермою. Потім соміти діляться на три частини: дорсолатеральну (дерматом), медіовентральну (склерот) і проміжну (міот). З появою соміт починається період розвитку зародка. На 21-й день у його складі виявляється 2-3 пари соміту; на 23-й день – 10 пар; на 25-й день – 14 пар; на 27-й день – 25 пар; наприкінці 5 тижнів - 43-44 пари.
Проміжки між зародковими листками та основними органами заповнюються зародковою сполучною тканиною - мезенхіма, яка складається з веретеноподібних або відрочастих клітин, що контактують відростками та занурені в аморфну міжклітинну речовину. Мезенхіма утворюється, головним чином, з мезодерми, однак у її розвитку беруть участь і інші зародкові листки, з яких ектодерма (М. Ф. Кащенко).
Коли вісцеральний листок вентральної мезодерми вступає в контакт із кишковою ентодермою, тіло зародка починає відмежовуватися від позазародкових органів тулубними складками. Цей процес починається з вентрального прогину країв зародка, який у результаті набуває дорсальної опуклості. Потім вигнуті краї зародка в головній та каудальній частинах починають заглиблюватися. Розширення цих відділяючих складок призводить до утворення бічних і черевних стінок зародка, а також виникнення так званого жовткового стеблинки, яке пов'язує його з жовтковим мішком. Із заглибленням тулубової складки кишкова ентодерма та вісцеральний листок вентральної мезодерми згортаються у трубку, що призводить до утворення кишки. Тим часом, як остання включає до свого складу кишкову ентодерму, жовтковий мішок містить жовткову ентодерму. Алантоїс, який раніше мав вигляд випинання жовткового мішка, тепер відходить від задньої кишки.
З розростанням амніотичної оболонки зростає обсяг амніону, який наближається до хоріону та зростається з ним. Амніотична ніжка при цьому переміщається з каудальної ділянки у вентральні стінки зародка і з'єднується з амніоном, утворюючи пупковий канатик (пуповину). Алантоїс, що раніше проник в амніотичну ніжку, частково включається до складу пуповини. В процесі цих перетворень жовтковий мішок віддаляється від зародка, а його зв'язок з кишечником зберігається завдяки трубчастій жовтково-кишковій протоці. Останній поряд з алантоїсом також знаходиться у складі пупкового канатика. Згодом алантоїс та жовтково-кишкова протока, що входять до складу пуповини, редукуються.
 Стадії ембріогенезу I – запліднення та утворення зиготи; II – дроблення та утворення бластули (бластоцисти); III – гаструляція – утворення зародкових листків та комплексу осьових органів; IV – гістогенез та органогенез зародкових та позазародкових органів; V - Системогенез.
Стадії ембріогенезу I – запліднення та утворення зиготи; II – дроблення та утворення бластули (бластоцисти); III – гаструляція – утворення зародкових листків та комплексу осьових органів; IV – гістогенез та органогенез зародкових та позазародкових органів; V - Системогенез.
 Пренатальний період починається з моменту запліднення та утворення зиготи та триває 280 днів. Ембріогенез поділяється на: - Початковий період (перший тиждень) - Ембріональний період (з 2-ї по 9 тиж.) - Плідний період (з 8 -9 тиж. до народження).
Пренатальний період починається з моменту запліднення та утворення зиготи та триває 280 днів. Ембріогенез поділяється на: - Початковий період (перший тиждень) - Ембріональний період (з 2-ї по 9 тиж.) - Плідний період (з 8 -9 тиж. до народження).

 ГАМЕТОГЕНЕЗ ✓ Первинні статеві клітини гоноцити (гонобласти) вперше виявляються у стінці жовткового мішка (наприкінці третього тижня життя ембріона людини). ✓ Гоноцити є індиферентними клітинами: вони не поділяються на чоловічі та жіночі клітини. ✓ Гоноцити мігрують у закладки статевих залоз та диференціюються у сперматогонії (у сім'янику) або в овонії (у яєчнику). Цей процес починається на 6-му тижні життя ембріона.
ГАМЕТОГЕНЕЗ ✓ Первинні статеві клітини гоноцити (гонобласти) вперше виявляються у стінці жовткового мішка (наприкінці третього тижня життя ембріона людини). ✓ Гоноцити є індиферентними клітинами: вони не поділяються на чоловічі та жіночі клітини. ✓ Гоноцити мігрують у закладки статевих залоз та диференціюються у сперматогонії (у сім'янику) або в овонії (у яєчнику). Цей процес починається на 6-му тижні життя ембріона.
 СПЕРМАТОГЕНЕЗ відбувається у звивистих канальцях насінника. У процесі сперматогенезу виділяють 4 фази (періоду): розмноження (проліферація); зростання; дозрівання; формування (сперміогенез).
СПЕРМАТОГЕНЕЗ відбувається у звивистих канальцях насінника. У процесі сперматогенезу виділяють 4 фази (періоду): розмноження (проліферація); зростання; дозрівання; формування (сперміогенез).
 I. Фаза розмноження: Чоловічі статеві клітини представлені диплоїдними клітинами – СПЕРМАТОГОНІЯМИ, які діляться мітотично. Розрізняють 3 типи сперматогоній (рис. 1): темні А сперматогонії – справжні стовбурові клітини, які стійкі до дії шкідливих факторів, діляться рідко; світлі А сперматогонії – напівстволові клітини, здатні до частих мітотичних поділів (результат поділу: утворюються дві А- сперматогонії, або одна А та одна В-). світлі У сперматогонії, комітовані клітини готуються до мейозу і вступають у фазу росту.
I. Фаза розмноження: Чоловічі статеві клітини представлені диплоїдними клітинами – СПЕРМАТОГОНІЯМИ, які діляться мітотично. Розрізняють 3 типи сперматогоній (рис. 1): темні А сперматогонії – справжні стовбурові клітини, які стійкі до дії шкідливих факторів, діляться рідко; світлі А сперматогонії – напівстволові клітини, здатні до частих мітотичних поділів (результат поділу: утворюються дві А- сперматогонії, або одна А та одна В-). світлі У сперматогонії, комітовані клітини готуються до мейозу і вступають у фазу росту.
 Ділянка стінки звивистого канальця сім'яника 1 - міоїдні клітини стінки; 2 – базальна мембрана; 3 – сперматогонії А; 4 – сперматогонії; 5 – сперматоцит першого порядку; 6 – сперматоцит другого порядку; 7 – сперматиди на ранній стадії розвитку; 8 – сперматиди наприкінці розвитку; 9 – клітина Сертолі.
Ділянка стінки звивистого канальця сім'яника 1 - міоїдні клітини стінки; 2 – базальна мембрана; 3 – сперматогонії А; 4 – сперматогонії; 5 – сперматоцит першого порядку; 6 – сперматоцит другого порядку; 7 – сперматиди на ранній стадії розвитку; 8 – сперматиди наприкінці розвитку; 9 – клітина Сертолі.
 ІІ. Фаза росту: У сперматогонії збільшуються в об'ємі (в 4 рази) і вступають у профазу першого мейотичного поділу, переміщаючись з базального шару у напрямку просвіту канальця → СПЕРМАТОЦИТИ 1-го порядку (диплоїдні).
ІІ. Фаза росту: У сперматогонії збільшуються в об'ємі (в 4 рази) і вступають у профазу першого мейотичного поділу, переміщаючись з базального шару у напрямку просвіту канальця → СПЕРМАТОЦИТИ 1-го порядку (диплоїдні).
 ІІІ. Фаза дозрівання: В результаті 1-го поділу мейозу (редукційного) утворюються СПЕРМАТОЦИТИ 2-го порядку, які швидко вступають у 2-е поділ мейозу (екваційне), клітини, що утворюються, називаються СПЕРМАТИДИ (гаплоїдні). Починаючи зі сперматогоній типу В, дочірні клітини при розподілі не відокремлюються повністю одна від одної, а залишаються пов'язаними за допомогою цитоплазматичних містків, утворюючи синцитії. Лише зрілі сперматозоїди відокремлюються від синцитію.
ІІІ. Фаза дозрівання: В результаті 1-го поділу мейозу (редукційного) утворюються СПЕРМАТОЦИТИ 2-го порядку, які швидко вступають у 2-е поділ мейозу (екваційне), клітини, що утворюються, називаються СПЕРМАТИДИ (гаплоїдні). Починаючи зі сперматогоній типу В, дочірні клітини при розподілі не відокремлюються повністю одна від одної, а залишаються пов'язаними за допомогою цитоплазматичних містків, утворюючи синцитії. Лише зрілі сперматозоїди відокремлюються від синцитію.

 IV. Фаза формування (сперміогенез): близько 50 діб. У процесі формування зрілих сперматозоїдів із сперматид відбуваються: конденсація ядерного хроматину; конденсація хроматину призводить до утворення суперкомпактного ядра, що збільшує мобільність сперматозоїда та захищає геном від ушкодження; освіта акросоми з елементів комплексу Гольджі; акросома - спеціальний тип лізосоми, що містить гідролітичні ферменти (такі як гіалуронідаза тощо) для руйнування оболонок яйцеклітини; центріолі переміщаються до протилежного полюса ядра; від дистальної центріолі утворюється осьова нитка (аксонема) джгутика. мітохондрії у вигляді спіралі оточують частину аксонеми, утворюючи потовщену ділянку - проміжний відділ; мітохондрії забезпечують енергію для руху сперматозоїда; цитоплазма редукується до мінімуму; після скидання надлишок цитоплазми, сперматозоїди відокремлюються від загальної асоціації клітин – синцитію – і стають вільними.
IV. Фаза формування (сперміогенез): близько 50 діб. У процесі формування зрілих сперматозоїдів із сперматид відбуваються: конденсація ядерного хроматину; конденсація хроматину призводить до утворення суперкомпактного ядра, що збільшує мобільність сперматозоїда та захищає геном від ушкодження; освіта акросоми з елементів комплексу Гольджі; акросома - спеціальний тип лізосоми, що містить гідролітичні ферменти (такі як гіалуронідаза тощо) для руйнування оболонок яйцеклітини; центріолі переміщаються до протилежного полюса ядра; від дистальної центріолі утворюється осьова нитка (аксонема) джгутика. мітохондрії у вигляді спіралі оточують частину аксонеми, утворюючи потовщену ділянку - проміжний відділ; мітохондрії забезпечують енергію для руху сперматозоїда; цитоплазма редукується до мінімуму; після скидання надлишок цитоплазми, сперматозоїди відокремлюються від загальної асоціації клітин – синцитію – і стають вільними.


 I. Фаза розмноження: На відміну від сперматогенезу фаза розмноження в овогенезі протікає виключно в ембріональний період розвитку. Овогонії діляться шляхом мітозів. Більшість дочірніх клітин, що утворюються, гине (цей процес називається атрезія), частина вступає у фазу росту (у профазу першого поділу мейозу). Мітотичні поділи вогонь припиняються до 7 місяця ембріогенезу. При народженні яйцеклітини в яєчнику є овоцитами першого порядку (первинними овоцитами).
I. Фаза розмноження: На відміну від сперматогенезу фаза розмноження в овогенезі протікає виключно в ембріональний період розвитку. Овогонії діляться шляхом мітозів. Більшість дочірніх клітин, що утворюються, гине (цей процес називається атрезія), частина вступає у фазу росту (у профазу першого поділу мейозу). Мітотичні поділи вогонь припиняються до 7 місяця ембріогенезу. При народженні яйцеклітини в яєчнику є овоцитами першого порядку (первинними овоцитами).
 ІІ. Фаза зростання: Фаза зростання відповідає життя первинного овоцита, що у профазі першого поділу мейозу, ділиться на два періоди: малого (до статевого дозрівання) і великого зростання. На момент народження в яєчнику налічується близько 2 млн первинних овоцитів; на період статевого дозрівання більшість їх гине, залишається близько 400 тисяч яйцеклітин. Зростання овоцитів відбувається у фолікулах – структурах, у яких овоцит оточений епітеліальними фолікулярними клітинами. У період малого зростання є два типи фолікулів: примордіальні фолікули (овоцит оточений одним шаром сплощених фолікулярних клітин) та первинні фолікули (з'являється блискуча (прозора оболонка), фолікулярні клітини мають кубічну або призматичну форму). Вступ овоцитів у період великого зростання відбувається лише під впливом гормонів (фолікуло-стимулюючого гормону); цьому періоду оогенезу відповідають зростаючі (вторинні та третинні) фолікули. Період зростання може тривати від 12 до 50 років.
ІІ. Фаза зростання: Фаза зростання відповідає життя первинного овоцита, що у профазі першого поділу мейозу, ділиться на два періоди: малого (до статевого дозрівання) і великого зростання. На момент народження в яєчнику налічується близько 2 млн первинних овоцитів; на період статевого дозрівання більшість їх гине, залишається близько 400 тисяч яйцеклітин. Зростання овоцитів відбувається у фолікулах – структурах, у яких овоцит оточений епітеліальними фолікулярними клітинами. У період малого зростання є два типи фолікулів: примордіальні фолікули (овоцит оточений одним шаром сплощених фолікулярних клітин) та первинні фолікули (з'являється блискуча (прозора оболонка), фолікулярні клітини мають кубічну або призматичну форму). Вступ овоцитів у період великого зростання відбувається лише під впливом гормонів (фолікуло-стимулюючого гормону); цьому періоду оогенезу відповідають зростаючі (вторинні та третинні) фолікули. Період зростання може тривати від 12 до 50 років.
 ІІІ. Фаза дозрівання Перший поділ мейозу завершується лише перед овуляцією (овуляція - розрив зрілого третинного фолікула та викид яйцеклітини з яєчника в черевну порожнину). Внаслідок мейозу I утворюється овоцит другого порядку (вторинний овоцит) – гаплоїдна клітина. Овоцит другого порядку входить у друге розподіл мейозу, але це розподіл блокується на стадії метафази і завершується лише за умови запліднення. За один менструально-оваріальний цикл (~ 28 днів) відбувається овуляція зазвичай однієї яйцеклітини. Таким чином, за репродуктивний період життя жінки (~3040 років) лише близько 450 яйцеклітин досягають зрілості – готовності до запліднення.
ІІІ. Фаза дозрівання Перший поділ мейозу завершується лише перед овуляцією (овуляція - розрив зрілого третинного фолікула та викид яйцеклітини з яєчника в черевну порожнину). Внаслідок мейозу I утворюється овоцит другого порядку (вторинний овоцит) – гаплоїдна клітина. Овоцит другого порядку входить у друге розподіл мейозу, але це розподіл блокується на стадії метафази і завершується лише за умови запліднення. За один менструально-оваріальний цикл (~ 28 днів) відбувається овуляція зазвичай однієї яйцеклітини. Таким чином, за репродуктивний період життя жінки (~3040 років) лише близько 450 яйцеклітин досягають зрілості – готовності до запліднення.
 Мейотичний поділ в овогенезі відрізняється тим, що протікає з нерівною цитотомією: у мейозі I хромосоми порівну розподіляються між дочірніми клітинами, але тільки одна з клітин (вторинний овоцит) зберігає майже всю цитоплазму; друга клітина (перше полярне, або редукційне, направне тільце) має дуже дрібні розміри, невелике ядро, мінімум цитоплазми.
Мейотичний поділ в овогенезі відрізняється тим, що протікає з нерівною цитотомією: у мейозі I хромосоми порівну розподіляються між дочірніми клітинами, але тільки одна з клітин (вторинний овоцит) зберігає майже всю цитоплазму; друга клітина (перше полярне, або редукційне, направне тільце) має дуже дрібні розміри, невелике ядро, мінімум цитоплазми.


 Яйцеклітина людини є вдруге оліголецитальною та ізолецитальною: у цитоплазмі рівномірно розподілено відносно невелику кількість жовтка; причому в еволюції це вдруге: вперше такий тип яйцеклітини зустрічається у ланцентника.
Яйцеклітина людини є вдруге оліголецитальною та ізолецитальною: у цитоплазмі рівномірно розподілено відносно невелику кількість жовтка; причому в еволюції це вдруге: вперше такий тип яйцеклітини зустрічається у ланцентника.
 Специфічні структури цитоплазми яйцеклітини. Деякі з цих речовин утворюються в печінці жінки, інші - безпосередньо в ооциті I. КОРТИКАЛЬНІ ГРАНУЛИ - Перебуваючи під плазмолемою, ці гранули містять ферменти, які після запліднення беруть участь у кортикальній реакції
Специфічні структури цитоплазми яйцеклітини. Деякі з цих речовин утворюються в печінці жінки, інші - безпосередньо в ооциті I. КОРТИКАЛЬНІ ГРАНУЛИ - Перебуваючи під плазмолемою, ці гранули містять ферменти, які після запліднення беруть участь у кортикальній реакції
 Особливості складу цитоплазми яйцеклітини У цитоплазмі - дуже високий вміст компонентів системи білоксинтезу (рибосом, т. РНК, м. РНК). ✓Відсутні центріолі - у зв'язку з цим здатність до поділів відновлюється тільки тоді, коли в клітину потрапляють центріолі сперматозоїда. ✓На поверхні плазмолеми є мікроворсинки. ✓Яйцеклітину (точніше, ооцит II), як і попередні клітини, оточують оболонки: блискуча, або прозора (zona pellucida, або Zp), і зерниста, утворена фолікулярними клітинами.
Особливості складу цитоплазми яйцеклітини У цитоплазмі - дуже високий вміст компонентів системи білоксинтезу (рибосом, т. РНК, м. РНК). ✓Відсутні центріолі - у зв'язку з цим здатність до поділів відновлюється тільки тоді, коли в клітину потрапляють центріолі сперматозоїда. ✓На поверхні плазмолеми є мікроворсинки. ✓Яйцеклітину (точніше, ооцит II), як і попередні клітини, оточують оболонки: блискуча, або прозора (zona pellucida, або Zp), і зерниста, утворена фолікулярними клітинами.
 Блискуча оболонка Zona pellucida: складається з глікопротеїнів різних видів (Zp 1, Zp 2, Zp 3) та глікозамінгліканів. Глікопротеїни фракції Zp 3 є рецепторами для сперматозоїдів, а глікопротеїни фракції Zp 2 після кортикальної реакції перешкоджають поліспермії. Компоненти блискучої оболонки синтезуються фолікулярними клітинами. Зерниста оболонка: фолікулярні клітини зернистої оболонки є частиною фолікулярного епітелію, мають довгі відростки, що пронизують блискучу оболонку. Променистий вінець: під променистим вінцем мають на увазі тільки внутрішні частини фолікулярних клітин з відростками, що відходять від них, або всю сукупність зернистого шару і блискучої оболонки. Редукційні тільця: десь у складі оболонок або під ними знаходяться редукційні тільця – інші (крім яйцеклітини) продукти двох поділів мейозу.
Блискуча оболонка Zona pellucida: складається з глікопротеїнів різних видів (Zp 1, Zp 2, Zp 3) та глікозамінгліканів. Глікопротеїни фракції Zp 3 є рецепторами для сперматозоїдів, а глікопротеїни фракції Zp 2 після кортикальної реакції перешкоджають поліспермії. Компоненти блискучої оболонки синтезуються фолікулярними клітинами. Зерниста оболонка: фолікулярні клітини зернистої оболонки є частиною фолікулярного епітелію, мають довгі відростки, що пронизують блискучу оболонку. Променистий вінець: під променистим вінцем мають на увазі тільки внутрішні частини фолікулярних клітин з відростками, що відходять від них, або всю сукупність зернистого шару і блискучої оболонки. Редукційні тільця: десь у складі оболонок або під ними знаходяться редукційні тільця – інші (крім яйцеклітини) продукти двох поділів мейозу.


 Головка Плазматична мембрана: Містить спеціальні білки, що беруть участь у таксі сперсматозоїда та зв'язуванні з яйцеклітиною ці білки (ті, що беруть участь у зв'язуванні з яйцеклітиною) мають високу видову специфічність. Акросома: Акросома - це сплощений мембранний мішечок, який подвійною шапочкою покриває ядро. Там містяться літичні ферменти (акрозин, гіалуронідаза та ін), що руйнують оболонки яйцеклітини. Ядро: різко ущільнене та містить гаплоїдний набір хромосом.
Головка Плазматична мембрана: Містить спеціальні білки, що беруть участь у таксі сперсматозоїда та зв'язуванні з яйцеклітиною ці білки (ті, що беруть участь у зв'язуванні з яйцеклітиною) мають високу видову специфічність. Акросома: Акросома - це сплощений мембранний мішечок, який подвійною шапочкою покриває ядро. Там містяться літичні ферменти (акрозин, гіалуронідаза та ін), що руйнують оболонки яйцеклітини. Ядро: різко ущільнене та містить гаплоїдний набір хромосом.
 Хвіст Шийка або сполучний відділ: Містить 2 центріолі. Від однієї з них починається аксонема, або осьова нитка хвоста, утворена мікротрубочками за схемою (9х2) + 2. Проміжна частина: У цій частині навколо аксонеми 9 зовнішніх фібрил, спіральна мітохондріальна оболонка і плазмолема. Головна або основна частина: Тут навколо аксонеми – фібрилярна піхва (9 зовнішніх фібрил та волокниста оболонка), а також плазмолема. Кінцева частина: У цій частині навколо аксонеми залишається лише плазмолема.
Хвіст Шийка або сполучний відділ: Містить 2 центріолі. Від однієї з них починається аксонема, або осьова нитка хвоста, утворена мікротрубочками за схемою (9х2) + 2. Проміжна частина: У цій частині навколо аксонеми 9 зовнішніх фібрил, спіральна мітохондріальна оболонка і плазмолема. Головна або основна частина: Тут навколо аксонеми – фібрилярна піхва (9 зовнішніх фібрил та волокниста оболонка), а також плазмолема. Кінцева частина: У цій частині навколо аксонеми залишається лише плазмолема.
 Еякулят чоловіка має об'єм 2 -3 мл і містить 200 -350 млн сперматозоїдів. У людини головка сперматозоїда сильно сплощена.
Еякулят чоловіка має об'єм 2 -3 мл і містить 200 -350 млн сперматозоїдів. У людини головка сперматозоїда сильно сплощена.
 Клітинний склад еякуляту людини А - зрілі, Б - незрілі 1, 2 - типовий сперматозоїд 3 -12 - атипові сперматозоїди 6 - 7 - аномалія форми головки та акросоми 8 -9 - аномалія джгутика 13 - 19 - незрілі 20 - 22 - лейкоцити
Клітинний склад еякуляту людини А - зрілі, Б - незрілі 1, 2 - типовий сперматозоїд 3 -12 - атипові сперматозоїди 6 - 7 - аномалія форми головки та акросоми 8 -9 - аномалія джгутика 13 - 19 - незрілі 20 - 22 - лейкоцити
 Сперматозоїди (на відміну від яйцеклітин) неоднорідні за видом статевої хромосоми, що міститься в їх ядрі: у 50% Сз є Х-хромосома, а в інших 50% Сз - Y-хромосома Стать дитини визначається "статтю" сперматозоїда:
Сперматозоїди (на відміну від яйцеклітин) неоднорідні за видом статевої хромосоми, що міститься в їх ядрі: у 50% Сз є Х-хромосома, а в інших 50% Сз - Y-хромосома Стать дитини визначається "статтю" сперматозоїда:
 Оптимальний термін для запліднення - перші 24 години після овуляції (хоча ооцит II може зберігати здатність до запліднення ще деякий час). Запліднення в нормі відбувається в ампулярній частині маткової труби
Оптимальний термін для запліднення - перші 24 години після овуляції (хоча ооцит II може зберігати здатність до запліднення ще деякий час). Запліднення в нормі відбувається в ампулярній частині маткової труби
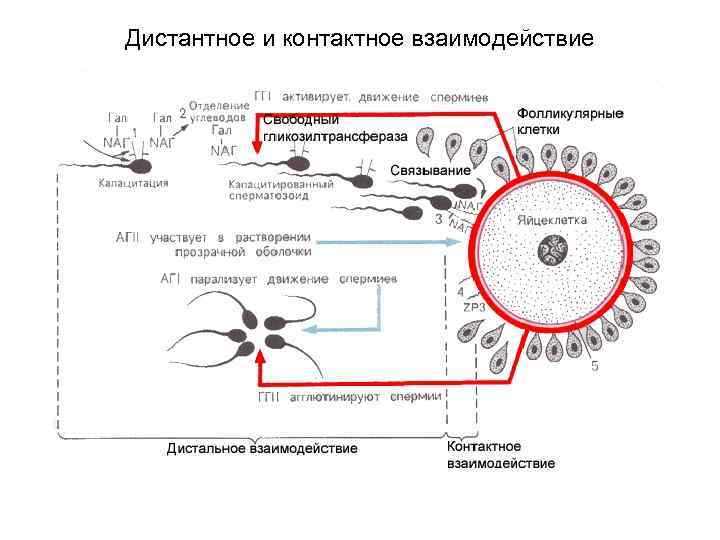
 Фази запліднення 1. Дистантна взаємодія та зближення гамет; 2. Контактна взаємодія та активізація яйцеклітини; 3. Входження сперматозоїда в яйцеклітину та подальше злиття – сингамія.
Фази запліднення 1. Дистантна взаємодія та зближення гамет; 2. Контактна взаємодія та активізація яйцеклітини; 3. Входження сперматозоїда в яйцеклітину та подальше злиття – сингамія.
 Запліднення 1, 2, 3, 4 – стадії акросомальної реакції 5 – блискуча зона 6 – перивителіновий простір 7 – плазматична мембрана 8 – кортикальна гранула 8 а – кортикальна реакція 9 – входження спермію в яйцеклітину 10 – зонна
Запліднення 1, 2, 3, 4 – стадії акросомальної реакції 5 – блискуча зона 6 – перивителіновий простір 7 – плазматична мембрана 8 – кортикальна гранула 8 а – кортикальна реакція 9 – входження спермію в яйцеклітину 10 – зонна
 I. Зближення та дистантна взаємодія статевих клітин Ооцит II повільно переміщається від яєчника у напрямку до матки; це відбувається пасивно – завдяки струму слизу. Даний струм викликається биттям вій миготливих клітин та тонічними скороченнями маткових труб (під дією прогестерону). У піхву власна рухливість сперматозоїдів (Сз) невелика через кисле середовище. У матку вони потрапляють, переважно, пасивно - завдяки тонічним скороченням жіночих статевих шляхів. Потім частина Сз також, переважно, пасивно, досягає маткових труб. Капацитація т. е. активація Сз: метаболізм і рухливість Сз різко посилюються, а мембрани Сз в ділянці головки втрачають поверхневі глікопротеїни і тому набувають здатності зв'язуватися з блискучою оболонкою ооцита Капацитацію ініціюють гіногамони II, що виділяються ооцитом. Активний рух Сз (реотаксис, хемотаксис) Рух Сз стає переважно активним і забезпечується биттям їх джгутиків. При цьому сперматозоїди одночасно рухаються поступально і обертаються навколо осі;
I. Зближення та дистантна взаємодія статевих клітин Ооцит II повільно переміщається від яєчника у напрямку до матки; це відбувається пасивно – завдяки струму слизу. Даний струм викликається биттям вій миготливих клітин та тонічними скороченнями маткових труб (під дією прогестерону). У піхву власна рухливість сперматозоїдів (Сз) невелика через кисле середовище. У матку вони потрапляють, переважно, пасивно - завдяки тонічним скороченням жіночих статевих шляхів. Потім частина Сз також, переважно, пасивно, досягає маткових труб. Капацитація т. е. активація Сз: метаболізм і рухливість Сз різко посилюються, а мембрани Сз в ділянці головки втрачають поверхневі глікопротеїни і тому набувають здатності зв'язуватися з блискучою оболонкою ооцита Капацитацію ініціюють гіногамони II, що виділяються ооцитом. Активний рух Сз (реотаксис, хемотаксис) Рух Сз стає переважно активним і забезпечується биттям їх джгутиків. При цьому сперматозоїди одночасно рухаються поступально і обертаються навколо осі;
 ІІ. Контактна взаємодія статевих клітин Зв'язування клітин Досягаючи ооциту II, численні Сз зв'язуються з його оболонками (за рахунок взаємодії певних рецепторів). При цьому через биття джгутиків Сз ооцит починає обертатися навколо власної осі. Акросомна реакція У зв'язаних Сз розвивається акросомна реакція: розриваються передні ділянки плазмолеми і мембрани акросоми, чому вивільняються акросомальні ферменти: гіалуронідаза роз'єднує клітини зернистої оболонки, а трипсиноподібний фермент акрозин і ряд ферментів акрозин і ряд ферментів акрозин і ряд.
ІІ. Контактна взаємодія статевих клітин Зв'язування клітин Досягаючи ооциту II, численні Сз зв'язуються з його оболонками (за рахунок взаємодії певних рецепторів). При цьому через биття джгутиків Сз ооцит починає обертатися навколо власної осі. Акросомна реакція У зв'язаних Сз розвивається акросомна реакція: розриваються передні ділянки плазмолеми і мембрани акросоми, чому вивільняються акросомальні ферменти: гіалуронідаза роз'єднує клітини зернистої оболонки, а трипсиноподібний фермент акрозин і ряд ферментів акрозин і ряд ферментів акрозин і ряд.
 ІІІ. Проникнення сперматозоїда в ооцит Ооцит проникають ядро Сз і центріолі. Після цього в ооцит II протягом декількох секунд розвивається кортикальна реакція - викид вмісту кортикальних гранул за межі клітини, утворюється оболонка запліднення. Крім того, ооцитом виділяються гіногамони I, які викликають аглютинацію сперматозоїдів, що залишилися. Завершення мейозу Одночасно проходять стадії другого поділу мейозу (метафаза, анафаза, телофаза). Це збільшує кількість редукційних, або полярних, тілець під блискучою оболонкою.
ІІІ. Проникнення сперматозоїда в ооцит Ооцит проникають ядро Сз і центріолі. Після цього в ооцит II протягом декількох секунд розвивається кортикальна реакція - викид вмісту кортикальних гранул за межі клітини, утворюється оболонка запліднення. Крім того, ооцитом виділяються гіногамони I, які викликають аглютинацію сперматозоїдів, що залишилися. Завершення мейозу Одночасно проходять стадії другого поділу мейозу (метафаза, анафаза, телофаза). Це збільшує кількість редукційних, або полярних, тілець під блискучою оболонкою.
 Підготовка зиготи до дроблення Зближення ядер У зиготі, що утворюється, ядро Сз набухає (перетворюючись на чоловічий пронуклеус і зближується з жіночим пронуклеусом (зближені ядра називаються синкаріоном), але не зливається з ним. Подвоєння ДНК і центріолей Подвоюються молекули ДНК центріолі Ці процеси тривають близько доби.
Підготовка зиготи до дроблення Зближення ядер У зиготі, що утворюється, ядро Сз набухає (перетворюючись на чоловічий пронуклеус і зближується з жіночим пронуклеусом (зближені ядра називаються синкаріоном), але не зливається з ним. Подвоєння ДНК і центріолей Подвоюються молекули ДНК центріолі Ці процеси тривають близько доби.
 Початок першого мітотичного розподілу Утворення єдиної материнської зірки У першому мітотичному розподілі беруть участь два пронуклеуси, що так і не злилися: їх оболонки руйнуються, а хромосоми конденсуються і в метафазі утворюють єдину материнську зірку. Весь цей час триває повільне пасивне просування ооциту II, а потім зиготи, по матковій трубі до матки.
Початок першого мітотичного розподілу Утворення єдиної материнської зірки У першому мітотичному розподілі беруть участь два пронуклеуси, що так і не злилися: їх оболонки руйнуються, а хромосоми конденсуються і в метафазі утворюють єдину материнську зірку. Весь цей час триває повільне пасивне просування ооциту II, а потім зиготи, по матковій трубі до матки.
 Малюнок - зигота людини на стадії синкаріону. На малюнку навколо зиготи видно щільну оболонку запліднення (1); вона не має клітинної структури, тому що походить з блискучої оболонки. Усередині зиготи – два ядра-пронуклеуси (2) майже рівного об'єму. Отже, дуже дрібне та щільне ядро сперматозоїда в результаті деконденсації хромосом та набухання досягає розміру ядра яйцеклітини. У кожному з цих ядер - гаплоїдний набір хромосом, тобто по 23 хромосоми. Оскільки ядра вже стикаються (що є сигналом до початку мітозу), в них вже відбулося подвоєння ДНК, і хромосоми стали двороматидними.
Малюнок - зигота людини на стадії синкаріону. На малюнку навколо зиготи видно щільну оболонку запліднення (1); вона не має клітинної структури, тому що походить з блискучої оболонки. Усередині зиготи – два ядра-пронуклеуси (2) майже рівного об'єму. Отже, дуже дрібне та щільне ядро сперматозоїда в результаті деконденсації хромосом та набухання досягає розміру ядра яйцеклітини. У кожному з цих ядер - гаплоїдний набір хромосом, тобто по 23 хромосоми. Оскільки ядра вже стикаються (що є сигналом до початку мітозу), в них вже відбулося подвоєння ДНК, і хромосоми стали двороматидними.
 Дроблення Дзигування – повне (голобластичне), нерівномірне (2 типи бластомерів: дрібні, світлі і більші, темні), асинхронне, повільне. Площина першого дроблення проходить через редукційні тільця.
Дроблення Дзигування – повне (голобластичне), нерівномірне (2 типи бластомерів: дрібні, світлі і більші, темні), асинхронне, повільне. Площина першого дроблення проходить через редукційні тільця.

 Дроблення. Загальна характеристика Збереження оболонки запліднення Протягом 2-4 діб відбувається дроблення, тобто сукупність мітотичних поділів без періодів зростання дочірніх клітин. Зростання клітин утруднений тому, що навколо зародка зберігається щільна оболонка запліднення, яка перешкоджає і припливу поживних речовин ззовні (життєдіяльність підтримується рахунок витрачання резервів яйцеклітини); і збільшення розміру зародка. Зменшення розміру клітин У силу вищесказаного, утворюються все дрібніші клітини і загальний обсяг зародка не збільшується.
Дроблення. Загальна характеристика Збереження оболонки запліднення Протягом 2-4 діб відбувається дроблення, тобто сукупність мітотичних поділів без періодів зростання дочірніх клітин. Зростання клітин утруднений тому, що навколо зародка зберігається щільна оболонка запліднення, яка перешкоджає і припливу поживних речовин ззовні (життєдіяльність підтримується рахунок витрачання резервів яйцеклітини); і збільшення розміру зародка. Зменшення розміру клітин У силу вищесказаного, утворюються все дрібніші клітини і загальний обсяг зародка не збільшується.

 Утворення бластоцисти Через 4, 5 - 5 діб утворюється бластоциста -зародкова бульбашка, заповнена рідиною. Компоненти бластоцисти такі: Більш дрібні світлі бластоміри утворюють трофобласт. Трофобласт – одношарова стінка з дрібних світлих клітин (згодом з трофобласту розвивається позазародковий орган – хоріон). Більші темні – Ембріобласт, або внутрішня клітинна маса-скупчення великих темних бластомерів у вигляді вузлика на внутрішній поверхні трофобласта в одного з полюсів. Бластоцель – порожнина, заповнена рідиною.
Утворення бластоцисти Через 4, 5 - 5 діб утворюється бластоциста -зародкова бульбашка, заповнена рідиною. Компоненти бластоцисти такі: Більш дрібні світлі бластоміри утворюють трофобласт. Трофобласт – одношарова стінка з дрібних світлих клітин (згодом з трофобласту розвивається позазародковий орган – хоріон). Більші темні – Ембріобласт, або внутрішня клітинна маса-скупчення великих темних бластомерів у вигляді вузлика на внутрішній поверхні трофобласта в одного з полюсів. Бластоцель – порожнина, заповнена рідиною.


 Зростання бластоцисти Завдяки всмоктування трофобластом рідини з порожнини матки, об'єм бульбашки дещо збільшується. У трофобласті з'являються вирости, що поступово руйнують оболонку запліднення навколо зародка. В результаті за кілька годин до імплантації зародок втрачає цю оболонку. Після цього оболонка не заважає зародку збільшуватися у вигляді, і з цього часу мітотичні цикли клітин стають звичайними, т. е. включають фазу зростання. Тому надалі збільшення маси зародка відбувається набагато швидше.
Зростання бластоцисти Завдяки всмоктування трофобластом рідини з порожнини матки, об'єм бульбашки дещо збільшується. У трофобласті з'являються вирости, що поступово руйнують оболонку запліднення навколо зародка. В результаті за кілька годин до імплантації зародок втрачає цю оболонку. Після цього оболонка не заважає зародку збільшуватися у вигляді, і з цього часу мітотичні цикли клітин стають звичайними, т. е. включають фазу зростання. Тому надалі збільшення маси зародка відбувається набагато швидше.
 Бластоциста залишається вільною в порожнині матки до 67 дня е. нар. потім занурюється в слизовий секрет маткових залоз на поверхні ендометрію.
Бластоциста залишається вільною в порожнині матки до 67 дня е. нар. потім занурюється в слизовий секрет маткових залоз на поверхні ендометрію.
 Імплантація - процес проникнення зародка в слизову оболонку стінки матки (ендометрій) та встановлення тісних зв'язків з її кровоносними судинами - початок на 6-7 день. Поділяється на дві стадії - адгезію та інвазію Під дією маткового секрету блискуча оболонка розчиняється, і бластоциста прикріплюється до ендометрію (адгезія). Клітини трофобласта швидко розростаються, виділяють лізосомальні ферменти та руйнують тканини ендометрію. Зазвичай імплантація відбувається в ендометрії задньої стінки матки.
Імплантація - процес проникнення зародка в слизову оболонку стінки матки (ендометрій) та встановлення тісних зв'язків з її кровоносними судинами - початок на 6-7 день. Поділяється на дві стадії - адгезію та інвазію Під дією маткового секрету блискуча оболонка розчиняється, і бластоциста прикріплюється до ендометрію (адгезія). Клітини трофобласта швидко розростаються, виділяють лізосомальні ферменти та руйнують тканини ендометрію. Зазвичай імплантація відбувається в ендометрії задньої стінки матки.

 Трофобласт З початком імплантації, (як тільки трофобласт входить в контакт з епітелієм ендометрію), клітини трофобласта починають активно ділитися, дочірні клітини зливаються, утворюючи єдину цитоплазматичну масу, що містить безліч ядер, і не має клітинних кордонів, утворюючи симпласто-(синцитіо) (Зовнішній листок) - багатоядерний симпласт, що утворюється в результаті злиття великої кількості клітин. і цитотрофобласт - (внутрішній листок) (зберігає клітинну будову);
Трофобласт З початком імплантації, (як тільки трофобласт входить в контакт з епітелієм ендометрію), клітини трофобласта починають активно ділитися, дочірні клітини зливаються, утворюючи єдину цитоплазматичну масу, що містить безліч ядер, і не має клітинних кордонів, утворюючи симпласто-(синцитіо) (Зовнішній листок) - багатоядерний симпласт, що утворюється в результаті злиття великої кількості клітин. і цитотрофобласт - (внутрішній листок) (зберігає клітинну будову);

 Після імплантації симпластотрофобласт адсорбує продукти розпаду тканин – гістотрофний тип харчування (перші два тижні). Таким чином роль симпластотрофобласта полягає в тому, що: завдяки дії лізосомальних ферментів забезпечує інвазію зародка в стінку матки; він забезпечує харчування, необхідне зростання зародка у перші два тижні життя, абсорбуючи продукти розпаду тканин матки (гістотрофний тип харчування); виконує ендокринну функцію (стб - місце утворення гормонів, включаючи ХГ (гормон, що стимулює утворення прогестерону жовтим тілом - визначається в крові або сечі жінок, починаючи з 10-го дня вагітності, і є основою тесту на вагітність.).
Після імплантації симпластотрофобласт адсорбує продукти розпаду тканин – гістотрофний тип харчування (перші два тижні). Таким чином роль симпластотрофобласта полягає в тому, що: завдяки дії лізосомальних ферментів забезпечує інвазію зародка в стінку матки; він забезпечує харчування, необхідне зростання зародка у перші два тижні життя, абсорбуючи продукти розпаду тканин матки (гістотрофний тип харчування); виконує ендокринну функцію (стб - місце утворення гормонів, включаючи ХГ (гормон, що стимулює утворення прогестерону жовтим тілом - визначається в крові або сечі жінок, починаючи з 10-го дня вагітності, і є основою тесту на вагітність.).

 Після повного занурення зародка дефект в ендометрії спочатку заповнюється кров'ю, що згорнулася, потім заростає сполучною тканиною і до 12 -13 дня покривається епітелієм. Навколо зародка утворюються лакуни, що заповнюються материнською кров'ю. У клітинах строми ендометрію розвивається т.з. децидуальна реакція: розростаються судини, з'являється набряклість, клітини збільшуються в обсязі та накопичують глікоген та ліпіди.
Після повного занурення зародка дефект в ендометрії спочатку заповнюється кров'ю, що згорнулася, потім заростає сполучною тканиною і до 12 -13 дня покривається епітелієм. Навколо зародка утворюються лакуни, що заповнюються материнською кров'ю. У клітинах строми ендометрію розвивається т.з. децидуальна реакція: розростаються судини, з'являється набряклість, клітини збільшуються в обсязі та накопичують глікоген та ліпіди.
 Гаструляція (2-й тиждень) Гаструляція у людини проходить у дві фази. 1) розщеплення (деламінація) на 7-му добу, - одночасно з імплантацією, утворюється епібласт та гіпобласт; 2) імміграції на 14 - 15 добу, утворюється первинна смужка, головний вузлик. 3)Между цими двома фазами йде утворення позазародкових органів, необхідні успішного розвитку зародка.
Гаструляція (2-й тиждень) Гаструляція у людини проходить у дві фази. 1) розщеплення (деламінація) на 7-му добу, - одночасно з імплантацією, утворюється епібласт та гіпобласт; 2) імміграції на 14 - 15 добу, утворюється первинна смужка, головний вузлик. 3)Между цими двома фазами йде утворення позазародкових органів, необхідні успішного розвитку зародка.
 Гаструляція 1-а фаза - деламінація - на 7 день ембріогенезу. Ембріобласт розщеплюється на епібласт та гіпобласт – майбутня позазародкова (жовточна) ентодерма.
Гаструляція 1-а фаза - деламінація - на 7 день ембріогенезу. Ембріобласт розщеплюється на епібласт та гіпобласт – майбутня позазародкова (жовточна) ентодерма.

 Мезодерма З епібласту виселяються клітини, що дають початок позазародковій мезодермі. Позародкова мезодерма утворює губчасту структуру, утворену тяжами клітин. Поступово обростає стінки амніотичного та жовткового пухирця, формуючи амніотичну оболонку (зсередини вистелена позазародковою ектодермою, зовні – позазародковою мезодермою) та жовтковий мішок (зсередини вистелений позазародковою ентодермою). При підростанні мезодерми до трофобласту формується хоріональна оболонка (позародкова мезодерма + цито- та симпластотрофобласт) та невеликі порожнини зливаються в єдину порожнину хоріону (синонім – позазародковий загалом).
Мезодерма З епібласту виселяються клітини, що дають початок позазародковій мезодермі. Позародкова мезодерма утворює губчасту структуру, утворену тяжами клітин. Поступово обростає стінки амніотичного та жовткового пухирця, формуючи амніотичну оболонку (зсередини вистелена позазародковою ектодермою, зовні – позазародковою мезодермою) та жовтковий мішок (зсередини вистелений позазародковою ентодермою). При підростанні мезодерми до трофобласту формується хоріональна оболонка (позародкова мезодерма + цито- та симпластотрофобласт) та невеликі порожнини зливаються в єдину порожнину хоріону (синонім – позазародковий загалом).


 Утворення жовткового мішка. Розростання позазародкової мезодерми Клітини гіпобласта мігрують і стуляються, утворюючи стінку жовткового бульбашки. Епібласт – дно амніотичної бульбашки; Гіпобласт - дах жовткової бульбашки. Позародкова мезодерма (первинна мезенхіма) утворює губчасту структуру.
Утворення жовткового мішка. Розростання позазародкової мезодерми Клітини гіпобласта мігрують і стуляються, утворюючи стінку жовткового бульбашки. Епібласт – дно амніотичної бульбашки; Гіпобласт - дах жовткової бульбашки. Позародкова мезодерма (первинна мезенхіма) утворює губчасту структуру.
 Зародок пов'язаний з хоріоном, що формується через амніотичну ніжку, утворену позазародковою мезодермою. Позародкова мезодерма вростає у вирости трофобласта, і первинні ворсинки хоріону стають вторинними (на 12 -13 е добу).
Зародок пов'язаний з хоріоном, що формується через амніотичну ніжку, утворену позазародковою мезодермою. Позародкова мезодерма вростає у вирости трофобласта, і первинні ворсинки хоріону стають вторинними (на 12 -13 е добу).
 1 – зародковий епібласт. 2 – амніотична ектодерма. 3 - амніотична порожнина (між двома попередніми листками). 4 - жовтковий пухирець (утворюється з гіпобласта). Поверхні, що обростаються позазародковою мезодермою: 5 А - зовнішня поверхня амніотичної бульбашки; 5 Б – зовнішня поверхня жовткового мішка; 5 - внутрішня поверхня трофобласта. 6 - хоріон: це трофобласт разом з мезодермою, що підросла до нього. 7 - амніотична ніжка: утворюється із позазародкової мезодерми та пов'язує зародок з хоріоном.
1 – зародковий епібласт. 2 – амніотична ектодерма. 3 - амніотична порожнина (між двома попередніми листками). 4 - жовтковий пухирець (утворюється з гіпобласта). Поверхні, що обростаються позазародковою мезодермою: 5 А - зовнішня поверхня амніотичної бульбашки; 5 Б – зовнішня поверхня жовткового мішка; 5 - внутрішня поверхня трофобласта. 6 - хоріон: це трофобласт разом з мезодермою, що підросла до нього. 7 - амніотична ніжка: утворюється із позазародкової мезодерми та пов'язує зародок з хоріоном.

 Другий тиждень ембріогенезу називають періодом «двійок»: утворюються два шари – епібласт та гіпобласт, які складають зародковий диск; розвиваються дві бульбашки – амніотичний та жовтковий; диференціюються два шари трофобласту - цито-і симпластотрофобласт. До кінця цього тижня трофобласт утворює розгалужені випинання - первинні ворсини, після чого називається вже хоріоном (ворсинчастою оболонкою).
Другий тиждень ембріогенезу називають періодом «двійок»: утворюються два шари – епібласт та гіпобласт, які складають зародковий диск; розвиваються дві бульбашки – амніотичний та жовтковий; диференціюються два шари трофобласту - цито-і симпластотрофобласт. До кінця цього тижня трофобласт утворює розгалужені випинання - первинні ворсини, після чого називається вже хоріоном (ворсинчастою оболонкою).

 Таким чином, наприкінці першого тижня – початку другого тижня пренатального розвитку відбуваються такі дуже важливі процеси, що збігаються за часом їх протікання, як: імплантація; перша фаза гаструляції; поява позазародкової мезодерми та розвиток позазародкових органів: амніону, жовткового мішка, хоріону. У зв'язку з цим, кінець першого тижня життя ембріона відноситься до критичних (визначальним) періодом розвитку (7-8 день е. р.)
Таким чином, наприкінці першого тижня – початку другого тижня пренатального розвитку відбуваються такі дуже важливі процеси, що збігаються за часом їх протікання, як: імплантація; перша фаза гаструляції; поява позазародкової мезодерми та розвиток позазародкових органів: амніону, жовткового мішка, хоріону. У зв'язку з цим, кінець першого тижня життя ембріона відноситься до критичних (визначальним) періодом розвитку (7-8 день е. р.)
 3-й тиждень розвитку Друга стадія гаструляції - імміграція відбувається на 14-15-й день ембріогенезу.
3-й тиждень розвитку Друга стадія гаструляції - імміграція відбувається на 14-15-й день ембріогенезу.

 Клітини, що іммігрують у зоні гензенівського вузлика, утворюють хордомезодермальний відросток, що росте під епібластом у напрямку до головного кінця по серединній лінії зародка. Клітини цього відростка диференціюються в головну мезодерму та хорду. Роль хорди у процесі розвитку: ✴хорда - вісь майбутнього хребта ✴хорда - первинний індуктор і організатор: під впливом сигнальних молекул клітин хорди відбувається диференціювання первинної ектодерми в нейроектодерму та утворення нервової платівки та нервової трубки. ✴nucleus pulposus
Клітини, що іммігрують у зоні гензенівського вузлика, утворюють хордомезодермальний відросток, що росте під епібластом у напрямку до головного кінця по серединній лінії зародка. Клітини цього відростка диференціюються в головну мезодерму та хорду. Роль хорди у процесі розвитку: ✴хорда - вісь майбутнього хребта ✴хорда - первинний індуктор і організатор: під впливом сигнальних молекул клітин хорди відбувається диференціювання первинної ектодерми в нейроектодерму та утворення нервової платівки та нервової трубки. ✴nucleus pulposus


 Мезенхіма Клітини мезенхіми виселяються зі всіх зародкових листків, найактивніше з вісцерального листка спланхнотома. З мезенхіми утворюються декілька типів тканин та структур: кров; всі типи сполучної тканини; гладка м'язова тканина, кровоносні судини та ендокард серця; мікроглі центральної нервової системи.
Мезенхіма Клітини мезенхіми виселяються зі всіх зародкових листків, найактивніше з вісцерального листка спланхнотома. З мезенхіми утворюються декілька типів тканин та структур: кров; всі типи сполучної тканини; гладка м'язова тканина, кровоносні судини та ендокард серця; мікроглі центральної нервової системи.


 Третинні ворсинки хоріону Кровоносні судини плода вростають у хоріональні ворсинки, які стають третинними (з'являються на 3-му тижні розвитку).
Третинні ворсинки хоріону Кровоносні судини плода вростають у хоріональні ворсинки, які стають третинними (з'являються на 3-му тижні розвитку).

 З 20-го дня - початок відокремлення тіла зародка від позазародкових оболонок - утворюється тулубна складка. Внаслідок утворення тулубових складок тіло зародка піднімається над провізорними органами та відокремлюється від них. При цьому зародок скручується в трубку. Одночасно це призводить до утворення з кишкової ентодерми кишкової трубки, яка відокремлюється від позазародкової ентодерми жовткового мішка.
З 20-го дня - початок відокремлення тіла зародка від позазародкових оболонок - утворюється тулубна складка. Внаслідок утворення тулубових складок тіло зародка піднімається над провізорними органами та відокремлюється від них. При цьому зародок скручується в трубку. Одночасно це призводить до утворення з кишкової ентодерми кишкової трубки, яка відокремлюється від позазародкової ентодерми жовткового мішка.

 Амніон утворює замкнуту порожнину навколо зародка. Функції: створення водного середовища певного хімічного складу та тиску для вільного розвитку, захист від механічних, гравітаційних стресів. Поступово порожнина розростається. До 7-го тижня розвитку амніотична мезодерма входить у контакт із мезодермою хоріону (амніо-хоріональна оболонка). Крім того, амніотичний епітелій обростає амніотичну ніжку.
Амніон утворює замкнуту порожнину навколо зародка. Функції: створення водного середовища певного хімічного складу та тиску для вільного розвитку, захист від механічних, гравітаційних стресів. Поступово порожнина розростається. До 7-го тижня розвитку амніотична мезодерма входить у контакт із мезодермою хоріону (амніо-хоріональна оболонка). Крім того, амніотичний епітелій обростає амніотичну ніжку.

 Амніон функціонує до народження (плодовий міхур). До кінця вагітності заповнений 1 -1, 5 л амніотичної рідини (навкоплодні води).
Амніон функціонує до народження (плодовий міхур). До кінця вагітності заповнений 1 -1, 5 л амніотичної рідини (навкоплодні води).
 Жовтковий мішок Функціонує тільки на ранніх стадіях розвитку: локалізація перших вогнищ кровотворення та утворення кровоносних судин та первинних статевих клітин. Після 7 -8 тижнів піддається регресії, залишається у вигляді тяжу клітин у пупковому канатиці, що спрямовує кровоносні судини до плаценти. На відміну від птахів, у людини (та інших ссавців) жовтковий мішок практично не містить жовтка, а заповнений серозною рідиною.
Жовтковий мішок Функціонує тільки на ранніх стадіях розвитку: локалізація перших вогнищ кровотворення та утворення кровоносних судин та первинних статевих клітин. Після 7 -8 тижнів піддається регресії, залишається у вигляді тяжу клітин у пупковому канатиці, що спрямовує кровоносні судини до плаценти. На відміну від птахів, у людини (та інших ссавців) жовтковий мішок практично не містить жовтка, а заповнений серозною рідиною.
 Алантоїс розвивається на 17 -18 добу у вигляді невеликого сліпого виросту вентральної стінки задньої кишки. Алантоїс вростає в амніотичну ніжку, у його стінці розвиваються пупкові кровоносні судини, які він підводить до хоріону (провідник судин). На 2-му місяці е. нар. редукується і разом із залишками жовткового мішка утворює клітинний тяж у складі пупкового канатика.
Алантоїс розвивається на 17 -18 добу у вигляді невеликого сліпого виросту вентральної стінки задньої кишки. Алантоїс вростає в амніотичну ніжку, у його стінці розвиваються пупкові кровоносні судини, які він підводить до хоріону (провідник судин). На 2-му місяці е. нар. редукується і разом із залишками жовткового мішка утворює клітинний тяж у складі пупкового канатика.
 Хоріон – плідна частина плаценти У формуванні хоріону розрізняють три періоди: передворсинчастий (7 -8 добу), період утворення ворсинок (до 50 доби), період котиледонів (з 50 по 90 добу). Утворений хоріональною пластиною (незародкова мезодерма) і виростами пластини - третинними ворсинками, що гілкуються, покриті трофобластом.
Хоріон – плідна частина плаценти У формуванні хоріону розрізняють три періоди: передворсинчастий (7 -8 добу), період утворення ворсинок (до 50 доби), період котиледонів (з 50 по 90 добу). Утворений хоріональною пластиною (незародкова мезодерма) і виростами пластини - третинними ворсинками, що гілкуються, покриті трофобластом.
 Формування первинних ворсин (кінець 2-го тижня) До середини 2-го тижня розвитку зародок вже повністю занурений в ендометрій, а трофобласт складається з двох шарів – зовнішнього (симпластотрофобласта) та внутрішнього (цитотрофобласта). До кінця цього тижня трофобласт утворює розгалужені випинання - первинні ворсини, після чого називається вже хоріоном (ворсинчастою оболонкою).
Формування первинних ворсин (кінець 2-го тижня) До середини 2-го тижня розвитку зародок вже повністю занурений в ендометрій, а трофобласт складається з двох шарів – зовнішнього (симпластотрофобласта) та внутрішнього (цитотрофобласта). До кінця цього тижня трофобласт утворює розгалужені випинання - первинні ворсини, після чого називається вже хоріоном (ворсинчастою оболонкою).
 Формування вторинних ворсин (початок 3-го тижня) На початку 3-го тижня у ворсини хоріону проникає позазародкова мезенхіма, що обростає зсередини всі поверхні, в т. ч. внутрішню поверхню трофобластахоріону. Тим самим формується сполучнотканинна строма ворсин, а самі ворсини стають вторинними.
Формування вторинних ворсин (початок 3-го тижня) На початку 3-го тижня у ворсини хоріону проникає позазародкова мезенхіма, що обростає зсередини всі поверхні, в т. ч. внутрішню поверхню трофобластахоріону. Тим самим формується сполучнотканинна строма ворсин, а самі ворсини стають вторинними.
 Формування третинних ворсин (кінець 3-го тижня) До кінця 3-го тижня усередині ворсин з клітин мезенхіми формуються первинні кровоносні судини, які побудовані як гемокапіляри і вступають у зв'язок із судинами мезодермальної платівки хоріону. Ворсини з розвиненими кровоносними судинами називаються третинними.
Формування третинних ворсин (кінець 3-го тижня) До кінця 3-го тижня усередині ворсин з клітин мезенхіми формуються первинні кровоносні судини, які побудовані як гемокапіляри і вступають у зв'язок із судинами мезодермальної платівки хоріону. Ворсини з розвиненими кровоносними судинами називаються третинними.
 Частина хоріона, що руйнує стінку матки і бере участь у освіті плаценти, формує складнорозгалужені ворсинки і зветься ворсинчастий хоріон. Решту поверхні складає гладкий хоріон. Найбільші ворсинки, що відходять від хоріональної пластини, звуться стовбурових ворсинок. Стовбурові ворсинки рясно гілкуються, найдрібніші гілочки носять назви термінальних ворсинок. Ворсинки, що впроваджуються в базальну платівку ендометрію, називаються якірними ворсинками. Зазвичай стовбурові ворсинки є якірними.
Частина хоріона, що руйнує стінку матки і бере участь у освіті плаценти, формує складнорозгалужені ворсинки і зветься ворсинчастий хоріон. Решту поверхні складає гладкий хоріон. Найбільші ворсинки, що відходять від хоріональної пластини, звуться стовбурових ворсинок. Стовбурові ворсинки рясно гілкуються, найдрібніші гілочки носять назви термінальних ворсинок. Ворсинки, що впроваджуються в базальну платівку ендометрію, називаються якірними ворсинками. Зазвичай стовбурові ворсинки є якірними.

 Плацента людини – єдиний орган, що складається з клітин двох генетично різних організмів. Складається з плодової частини (хорион з ворсинками) материнської частини – змінений ендометрій
Плацента людини – єдиний орган, що складається з клітин двох генетично різних організмів. Складається з плодової частини (хорион з ворсинками) материнської частини – змінений ендометрій
 Плацентація - період ембріогенезу, протягом якого відбувається розвиток плаценти, один із критичних періодів ембріогенезу, відповідає 3 -8 тижням вагітності. Поверхневий шар ендометрію отримав назву децидуальної (відпадаючої) оболонки. Залежно від розташування щодо місця імплантації розрізняють: Decidua parietalis (пристіночна) – ендометрій, що вистилає порожнину матки за винятком ділянки імплантації; Decidua capsularis (сумкова) – частина ендометрію, яка оточує ембріон, що розвивається, утворюючи поверх нього капсулу, і відокремлює зародок від порожнини матки (до 16-ї тиж.) Decidua basalis (основна), материнська частина плаценти, та частина ендометрію, яка знаходиться між плодом та базальним шаром ендометрію. Отже, материнська частина плаценти представлена: базальною платівкою (decidua basalis) ендометрію; лакунами, заповненими материнською кров'ю.
Плацентація - період ембріогенезу, протягом якого відбувається розвиток плаценти, один із критичних періодів ембріогенезу, відповідає 3 -8 тижням вагітності. Поверхневий шар ендометрію отримав назву децидуальної (відпадаючої) оболонки. Залежно від розташування щодо місця імплантації розрізняють: Decidua parietalis (пристіночна) – ендометрій, що вистилає порожнину матки за винятком ділянки імплантації; Decidua capsularis (сумкова) – частина ендометрію, яка оточує ембріон, що розвивається, утворюючи поверх нього капсулу, і відокремлює зародок від порожнини матки (до 16-ї тиж.) Decidua basalis (основна), материнська частина плаценти, та частина ендометрію, яка знаходиться між плодом та базальним шаром ендометрію. Отже, материнська частина плаценти представлена: базальною платівкою (decidua basalis) ендометрію; лакунами, заповненими материнською кров'ю.
 Котіледони Ворсинки, звернені до decidua basalis, розподілені нерівномірно, групами – котиледонами. Котиледон – структурно-функціональна одиниця сформованої плаценти. Він утворений стволовою ворсинкою та її розгалуженнями. Котиледони частково розділені сполучнотканинними септами (плацентарними перегородками).
Котіледони Ворсинки, звернені до decidua basalis, розподілені нерівномірно, групами – котиледонами. Котиледон – структурно-функціональна одиниця сформованої плаценти. Він утворений стволовою ворсинкою та її розгалуженнями. Котиледони частково розділені сполучнотканинними септами (плацентарними перегородками).

 Гематоплацентарний бар'єр Кров матері та дитини ніколи не змішується Компоненти гематоплацентарного бар'єру, що розділяє кров матері та кров плода: ендотелій капіляра плода; базальна мембрана у стінці капілярів плода; сполучна тканина ворсинок (з клітинами-макрофагами); базальна мембрана трофобласту; цитоторофобласт; симпластотрофобласт. Зв'язок між циркуляціями крові плода та матері здійснюється через пупковий канатик.
Гематоплацентарний бар'єр Кров матері та дитини ніколи не змішується Компоненти гематоплацентарного бар'єру, що розділяє кров матері та кров плода: ендотелій капіляра плода; базальна мембрана у стінці капілярів плода; сполучна тканина ворсинок (з клітинами-макрофагами); базальна мембрана трофобласту; цитоторофобласт; симпластотрофобласт. Зв'язок між циркуляціями крові плода та матері здійснюється через пупковий канатик.
 Гематоплацентарний бар'єр Кров матері (1) та дитини (8) не поєднується: - ендотелій капілярів плода (7); - базальна мембрана у стінці капілярів плода (6); - сполучна тканина ворсинок (5); - базальна мембрана трофобласту (4); - цитоторофобласт (3); - симпластотрофобласт (2)
Гематоплацентарний бар'єр Кров матері (1) та дитини (8) не поєднується: - ендотелій капілярів плода (7); - базальна мембрана у стінці капілярів плода (6); - сполучна тканина ворсинок (5); - базальна мембрана трофобласту (4); - цитоторофобласт (3); - симпластотрофобласт (2)

 Гематоплацентарний проникаємо для ряду речовин і збудників хвороб Збудники хвороб Віруси (ВІЛ, цитомегаловірус, краснухи, віспи, вітряної віспи, кору, поліомієліту) Бактерії (туберкульоз, Treponema) , дифенін тетрациклін протиракові препарати, анестетики, заспокійливі, анальгетики Антитіла до резус-фактору
Гематоплацентарний проникаємо для ряду речовин і збудників хвороб Збудники хвороб Віруси (ВІЛ, цитомегаловірус, краснухи, віспи, вітряної віспи, кору, поліомієліту) Бактерії (туберкульоз, Treponema) , дифенін тетрациклін протиракові препарати, анестетики, заспокійливі, анальгетики Антитіла до резус-фактору
 Функції плаценти трофічна (з організму матері до плоду надходять найрізноманітніші поживні речовини, електроліти, вітаміни, деякі гормони; дихальна (транспорт кисню в кров плода та перенесення вуглекислого газу в кров матері); видільна (з крові плода в кров матері надходять продукти метаболізму та виділяються через нирки матері), захисна (запобігає розвитку імунного конфлікту, проникненню лікарських речовин – бар'єр не абсолютний), ендокринна (синтез низки гормонів та інших біологічно активних речовин).
Функції плаценти трофічна (з організму матері до плоду надходять найрізноманітніші поживні речовини, електроліти, вітаміни, деякі гормони; дихальна (транспорт кисню в кров плода та перенесення вуглекислого газу в кров матері); видільна (з крові плода в кров матері надходять продукти метаболізму та виділяються через нирки матері), захисна (запобігає розвитку імунного конфлікту, проникненню лікарських речовин – бар'єр не абсолютний), ендокринна (синтез низки гормонів та інших біологічно активних речовин).
 Обмін через плацентарний бар'єр З материнської крові в кров плода З крові плода в материнську (постачання) кров (виділення) кисень поживні речовини (глюкоза, амінокислоти, вільні жирні кислоти, вітаміни) електроліти вода стероїдні гормони (кортизол, естроген) антитіла (I - забезпечують пасивний імунітет плоду) білки плазми крові (альбумін) деякі білкові гормони (тироксин, інсулін) еритроцити (дуже мало) вуглекислий газ метаболіти (сечовина, сечова кислота, білірубін, креатин, креатинін) еритроцити (дуже мало)
Обмін через плацентарний бар'єр З материнської крові в кров плода З крові плода в материнську (постачання) кров (виділення) кисень поживні речовини (глюкоза, амінокислоти, вільні жирні кислоти, вітаміни) електроліти вода стероїдні гормони (кортизол, естроген) антитіла (I - забезпечують пасивний імунітет плоду) білки плазми крові (альбумін) деякі білкові гормони (тироксин, інсулін) еритроцити (дуже мало) вуглекислий газ метаболіти (сечовина, сечова кислота, білірубін, креатин, креатинін) еритроцити (дуже мало)
 Типи плацент у ссавців Епітеліохоріальна (свині): ворсинки хоріону проникають у поглиблення слизової оболонки (маткові залози), не руйнуючи їх. Десмохоріальна плацента (копитні): ворсинки хоріону контактують із сполучною тканиною ендометрію матері. Ендотеліо-(вазо-)хоріальна (хижаки) плацента: ворсинки хоріону контактують зі стінкою судин ендометрію матері, не руйнуючи стінки. Гемохоріальна плацента: ворсинки хоріону руйнують епітелій, сполучну тканину, стінки судин матері та омиваються кров'ю, що заповнюють лакуни.
Типи плацент у ссавців Епітеліохоріальна (свині): ворсинки хоріону проникають у поглиблення слизової оболонки (маткові залози), не руйнуючи їх. Десмохоріальна плацента (копитні): ворсинки хоріону контактують із сполучною тканиною ендометрію матері. Ендотеліо-(вазо-)хоріальна (хижаки) плацента: ворсинки хоріону контактують зі стінкою судин ендометрію матері, не руйнуючи стінки. Гемохоріальна плацента: ворсинки хоріону руйнують епітелій, сполучну тканину, стінки судин матері та омиваються кров'ю, що заповнюють лакуни.
Запліднення
заплідненнямназивається процес злиття зрілих чоловічої (сперматозоїд) та жіночої (яйцеклітина) статевих клітин, внаслідок чого виникає зигота, що несе генетичну інформацію як батька, так і матері.
Процес дозрівання чоловічих та жіночих статевих клітин дуже складний. Сперматогенез відбувається у звивистих насіннєвих канальцях чоловічих гонад. Він завершується в період статевої зрілості утворенням зрілих сперматозоїдів, які мають здатність до запліднення. Повному дозріванню передує процес редукційного поділу, в результаті якого в сперматозоїдному ядрі міститься гаплоїдний набір хромосом.
Сперматозоїди бувають двох видів: носії статевих Х- та Y-хромосом. При злитті з яйцеклітиною сперматозоїда, що є носієм статевої Х-хромо-соми, з зиготи, що утворюється, розвивається ембріон жіночої статі, при злитті сперматозоїда, що має статеву Y-хромосому, виникає зародок чоловічої статі (яйцеклітина завжди є носієм).
Зрілий сперматозоїд має довжину до 50-60 мкм і складається з голівки, шийки та хвостової частини. Головка сперматозоїда, що має овальну форму, містить ядро, оточене тонким шаром протоплазми. Шийка має протоплазму, що містить видозмінену центросому, яка сприяє процесу дроблення заплідненої яйцеклітини. Хвостик складається з протоплазми та виконує рухові функції. В результаті коливальних рухів хвостової частини сперматозоїди здатні здійснювати самостійні рухи у статевому тракті жінки зі швидкістю 2-3 мм/хв. Здатність до рухів сперматозоїди отримують після їх контакту з секретом насіннєвих бульбашок та передміхурової залози. Така суміш носить назву насіннєвої рідини, або сперми. При статевих зносинах у піхву жінки виливається в середньому близько 3-5 мл сперми, в якій міститься 300-500 млн сперматозоїдів. Використання електронної мікроскопії дозволило встановити складну структуру сперматозоїда (рис. 3.1).
Насіннєва рідина має складний склад і містить фруктозу, білкові речовини, протеази, кислу фосфатазу, лимонну кислоту та біологічно активні речовини – простагландини. Насіннєва рідина має виражену антигенну активність, при цьому антигени виявлені як у самих сперматозоїдах, так і в рідкій частині сперми. Вважають, що кількість антигенних компонентів у насінній рідині досягає 12. У процесі запліднення відбувається нейтралізація сперматозоїдів антигенів.
Розвиток яйцеклітин пов'язане зі зростанням та розвитком первинних фолікулів, що знаходяться в кірковому шарі яєчників. Дозрівання яйцеклітини умовно можна розділити на дозрівання ядра та дозрівання цитоплазми. Під дозріванням ядра розуміють сукупність процесів, що переводять ядро зі стадії диплотена I (або стадії гермінативної бульбашки) до метафази другого мейотичного поділу. Дозрівання ядра не включає завершення мейозу, так як тільки проникнення сперматозоїда всередину яйцеклітини викликає завершення другого редукційного поділу.
Головка Тіло Хвіст
Рис.3.1. Будова сперматозоїда людини (електронно-мікроскопічна схема). 1 – акросома; 2 – ядро; 3 – шийка; 4 - мітохондрії; 5 – осьові нитки.

Рис. 3.2. Зріла яйцеклітина, оточена клітинами променистого вінця.
Від періоду настання статевого дозрівання до менопаузи у жінки у кожному менструальному циклі зазвичай дозріває одна яйцеклітина. Овоцит першого порядку перетворюється на овоцит другого порядку при відщепленні першого полярного тільця. У момент овуляції овоцит другого порядку виявляється блокованим на стадії мета-фази другого мейотичного поділу. Дозрівання яйцеклітин стало відоме в деталях завдяки можливості культивування яйцеклітин in vitro, що в даний час широко використовують при екстракорпоральному заплідненні.
Зріла яйцеклітина складається з ядра, цитоплазми, оточена блискучою оболонкою і клітинами променистого вінця, що є залишками гранульозних клітин фолікула (рис. 3.2). Жіноча статева клітина, як і чоловіча, має антигенні властивості. Особливо багата на різні антигени її блискуча оболонка.
Перш ніж аналізувати механізми запліднення, тобто. процес злиття яйцеклітини зі сперматозоїдом, слід висвітлити питання транспорту гамет. Насамперед це стосується просування сперматозоїдів каналами репродуктивної системи жінки. При нормальній еякуляції у піхву потрапляє в середньому близько 100 млн. сперматозоїдів, деякі з яких мають ті чи інші морфологічні або функціональні відхилення. Частина сперматозоїдів, у тому числі й неповноцінних, залишається у піхві та піддається фагоцитозу. Разом із сперматозоїдами у піхву потрапляють й інші складові сперми, при цьому особлива роль належить простагландинам. Під їх впливом відбувається активація скорочувальної активності матки та маткових труб, що дуже важливо для нормального транспортування гамет.
З піхви, що має у здорової жінки кисле середовище (кисле
середовище несприятливе для життєдіяльності сперматозоїдів), останні швидко надходять до цервікального слизу, який під час статевого акту під впливом скорочень м'язів шийки матки виділяється з цервікального каналу. Наявність слаболужної реакції цервікального слизу сприяє підвищенню рухової активності сперматозоїдів.
Оптимальний склад цервікального слизу формується на момент овуляції переважно під впливом естрогенних гормонів яєчників. У цей період міцелії слизу розташовуються у вигляді своєрідних ланцюжків, орієнтованих на силові лінії магнітного поля землі. Сперматозоїди просуваються по міцеліям слизу у напрямку до матки, але лише за певної архітектоніки ланцюжків міцел. Турбулентні рухи сперматозоїдів найбільш виражені в пристінкових областях шийки матки, при цьому частина сперматозоїдів на деякий час може депонуватися в криптах шийки матки, створюючи тим самим своєрідний резерв сперми, звідки надалі може відбуватися їх додаткове надходження вгору по каналах репродуктивної системи.
У верхніх відділах статевого тракту жінки починається процес, що називається капацитацією сперми, - набуття нею завдяки складним змінам здатності до запліднення. В результаті капацитації сперматозоїди набувають здатності до акросомальної реакції. Крім цього, капацитація виявляється у змінах рухів хвостових частин сперматозоїдів (наявність надактивної рухливості). Тонкі механізми капацитації досі повністю не вивчені. Час капацитації по-різному у різних сперматозоїдів, що, мабуть, є важливою пристосувальною реакцією для процесу запліднення. Капацитовані сперматозоїди дуже активні, проте тривалість їхнього життя менша, ніж некапацитованих. Капацитовані сперматозоїди мають підвищену здатність пенетрувати тканини, що має вирішальне значення в процесі запліднення яйцеклітини.
Транспорт сперматозоїдів до матки, а потім і до маткових труб в основному забезпечується скороченнями гладкої мускулатури цих органів. Вважають також, що трубно-маточні сфінктери є своєрідними дозаторами надходження сперматозоїдів із порожнини матки у просвіти маткових труб.
Поряд із скорочувальною здатністю маткових труб, які знаходяться під складним гормональним впливом (естрогени, андрогени, окситоцин), а також під впливом простагландинів, велике значення у просуванні сперматозоїдів, крім їх власної високої кінетичної активності, належить таким факторам, як рух мікроворсинянсендят та струм рідини у просвіті маткової труби. Таким чином, переміщення сперматозоїдів каналами репродуктивної системи жінки є надзвичайно складним багатокомпонентним процесом.
Результати сучасних наукових досліджень свідчать про наявність двофазності процесу транспорту сперматозоїдів маточними трубами. У першу (коротку) фазу сперматозоїди починають швидко надходити в ампулу труби. Ця фаза триває лише кілька хвилин і регулюється скоротливою активністю матки та маткових труб. Дана фаза змінюється тривалішою другою фазою, протягом якої сперматозоїди зі значно меншою швидкістю транспортуються до місця запліднення.
В ампулярній частині маткової труби нормальна кількість сперматозоїдів у тривалій фазі транспорту зберігається на певному рівні завдяки безперервному відшкодуванню сперматозоїдів, що йдуть у черевну порожнину за рахунок тих статевих клітин, які депоновані в нижніх відділах статевого тракту (крипти шийки матки та ін).
Велике значення у транспорті гамет та заплідненні має так зване захоплення яйцеклітини ампулярним відділом маткової труби. Цей процес полягає в тому, що ампулярний відділ труби своїми фімбріями як би накриває яєчник там, де відбулася овуляція; при цьому значно полегшується переміщення зрілої яйцеклітини в ампулу маткової труби. Механізм цього феномену ще остаточно не досліджений.
Істотним є питання про виживання сперматозоїдів у статевих шляхах жінки. Деякі автори вважають, що життєздатність сперми зберігається протягом кількох (до 5) днів. Однак слід враховувати, що збереження рухливості сперматозоїдів не обов'язково свідчить про їхню здатність, що запліднює.
А У найбільш сприятливих умовах, коли чоловічі статеві клітини знаходяться в цервікальному слизу на тлі високого вмісту естрогенів в організмі, здатність сперматозоїдів, що запліднює, тримається до 2 діб після еякуляції у піхву. У зв'язку з цим прийнято вважати, що для досягнення вагітності оптимальна частота статевих зносин у період до та після овуляції має бути кожні 2 дні. При частіших статевих зносинах фертильність сперми знижується.
Перед заплідненням зріла яйцеклітина з усіх боків оточена променистим вінцем (corona radiata). Виразно помітна блискуча оболонка (zona pellucida). Вже за кілька хвилин після розриву фолікула (овуляції) яйцеклітина потрапляє в порожнину маткової труби. Цьому процесу сприяє ряд факторів: "захоплення" її фимбріями маткової труби з боку яєчника, в якому відбулася овуляція, напрямок струму фолікулярної рідини при розриві фолікула та ін. Цим факторам належить дуже важлива роль у початковому транспорті яйцеклітини, яка позбавлена самостійної рухливості. Здатність яйцеклітини до запліднення в середньому становить 24 години. Сучасні методи діагностики (ультразвукове дослідження, лапароскопія) дозволяють не тільки спостерігати за процесом овуляції, а й фіксувати цей процес на фотоплівку.
Яйцеклітина, що потрапила до ампулярного відділу маткової труби, швидко оточується великою кількістю сперматозоїдів, що є носіями як Х-, так і Y-статевих хромосом. Під мікроскопом сперматозоїди з Х-хромосомою мають дещо більші розміри, ніж несучі Y-хромосому. Сперматозоїди, що оточують яйцеклітину, починають пенетрувати в клітини променистого вінця. Процес пенетрації обумовлений наявністю ряду ферментів, що містяться як у головці сперматозоїда, так і у трубній рідині (рис. 3.3).
Відразу після злиття мембран статевих клітин відбувається кортикальна реакція яйцеклітини, що є складовою забезпечення блоку поліспермії. Після цього хромосоми зиготи вступають у перший мітотичний поділ, який настає через 24 години після початку запліднення. Ядро заплідненої яйцеклітини (зиготи) містить диплоїд-

Рис. 3.3. Пенетрація сперматозоїдів у яйцеклітину в ампулярному відділі маткової труби (схема).
ний набір хромосом (46). Таким чином, новий організм є носієм генетичної інформації обох батьків.
Після запліднення (через 24 год) починається дроблення заплідненої яйцеклітини. Спочатку дроблення має синхронний характер. Через 12 год від початку виникнення 2 бластомерів виникають 4 бластомери і т.д. До 96 год від моменту злиття ядра сперматозоїда з ядром яйцеклітини зародок складається з 16-32 бластомерів (стадія морули). На цій стадії запліднене яйце (зигота) потрапляє до матки.
Оскільки яйцеклітина, що дробиться, не володіє самостійною рухливістю, її транспорт визначається взаємодією скорочувальної активності маткової труби (основний фактор), рухами циліарного епітелію ендосальпінксу і капілярним струмом рідини в напрямку від ампулярного кінця маткової труби до матки.
Транспорт яйцеклітини матковою трубою перебуває під впливом гормонів. Як відомо, після овуляції на місці фолікула, що лопнув, утворюється нова ендокринна залоза - жовте тіло. Воно виділяє як прогестерон, і естрогенні гормони. Саме цим двом статевим гормонам належить провідна роль забезпеченні кінетики маткових труб. Під впливом щодо низького вмісту прогестерону та вищої концентрації в крові естрогенів (що має місце безпосередньо після овуляції) підвищується тонус ампулярно-перешийкового відділу труби. В результаті яйцеклітина затримується в ампулярному відділі, де відбувається її запліднення та починається процес клітинного поділу з утворенням бластомерів. Надалі відбувається поступове просування заплідненої яйцеклітини по перешийку маткової труби до матки. Під впливом наростаючих концентрацій прогестерону жовтого тіла скорочувальна функція маткових труб набуває перистальтичного характеру, при цьому хвилі скорочень спрямовані у бік матки. Відбувається розслаблення трубно-маткової сполуки, і яйцеклітина з маткової труби потрапляє у порожнину матки.
 Ендометрій
Ендометрій
Рис. 3.4. Транспорт заплідненої яйцеклітини матковою трубою аж до імплантації (схема).
1 – яйцеклітина в ампулі маткової труби; 2 – запліднення; 3-7 – різні стадії утворення бластомерів; 8 – морула; 9, 10 – бластоциста; 11 – імплантація.
Аналізуючи механізми гормональної регуляції кінетики маткових труб, слід підкреслити, що лише певне співвідношення між концентраціями в крові прогестерону та естрогенів забезпечує описану вище чітку фізіологічну взаємодію різних відділів маткової труби та своєчасний транспорт плодового яйця в порожнину матки. Поряд із прогестероном та естрогенами у здійсненні фізіологічної кінетики маткових труб у ранні терміни вагітності відоме значення відводиться таким гормонам, як тестостерон та кортизол. Проте роль цих гормонів вивчена нині ще недостатньо.
Імплантація плодового яйця.Пройшовши шлях по матковій трубі протягом 4 діб, плодове яйце на стадії морули потрапляє в матку, де перетворюється на бластоцисту (рис. 3.4). Стадія бластоцисти характеризується тим, що блас-томери зазнають певних змін. Частина бластомерів, більших за своїми розмірами, утворює так званий ембріобласт,з якого надалі розвивається ембріон. Інша частина клітин, дрібніших і розташовуються по периферії плодового яйця, утворює поживну оболонку - трофобласт.Надалі найбільш розвинена частина трофобласту перетворюється на плаценту.У порожнині матки бластоциста наближається до місця імплантації (нідації) (рис. 3.5 а,б). Локалізація імплантації має свої закономірності і, мабуть, значною мірою визначається місцевими особливостями ендометрію. Зазвичай бластоциста імплантується в ділянці передньої або задньої стінки матки.
Потім починається занурення бластоцисти в ендометрій, який на той час перетворюється на децидуальну оболонку (рис. 3.5, в, г).
 |
 |
Рис. 3.5. Імплантація.
а – бластоциста перед імплантацією; б - початковий контакт бластоцисти з деци-дуальною оболонкою матки; в - занурення бластоцисти у децидуальну оболонку; г – завершення імплантації.
Децидуальна оболонка є видозміненим функціональним шаром ендометрію. До моменту імплантації заплідненої яйцеклітини слизова оболонка матки знаходиться в секреторній фазі, залози пилкоподібно змінені і заповнені секретом, клітини строми мають округлий вигляд і містять велику кількість глікогену, ліпідів, нейтральних мукополісахаридів, солей і мікроелементів йоні активні сполуки, необхідні життєдіяльності зародка.
Процес імплантації насамперед пов'язані з гормональними чинниками. Провідна роль належить статевим (стероїдним) гормонам. Під час вагітності відбувається розвиток і активне функціонування жовтого тіла яєчника, що виник на місці фолікула, що лопнув. Жовте тіло секретує велику кількість прогестерону та дещо менше – естрогенів. Ці статеві гормони через специфічні стероїдні рецептори, що знаходяться в ендометрії, надають виражений вплив на секреторні перетворення слизової оболонки матки та процеси її децидуалізації. Крім стероїдних, певну роль в імплантації відіграють і деякі інші гормони (пролактин, глюкокортикоїди).
Децидуальна тканина як своєрідний ендокринний орган має пряме відношення до імплантації та подальших стадій постімплантаційного розвитку зародка. Встановлено, що ендометрій жінки виробляє імунореактивний пролактин. Він починає секретуватися з 9-го дня після овуляції, при цьому під час вагітності продукція тканинного пролактину значно підвищується, тому нині деякі дослідники схильні розглядати децидуальну тканину як своєрідний ендокринний орган.
Процес імплантації, що у середньому триває близько 2 днів, супроводжується як значними змінами клітинних елементів залоз і строми ендометрію, а й вираженими гемодинамическими зрушеннями місцевого характеру. Поблизу місця імплантації бластоцисти відзначається розширення кровоносних судин та утворення синусоїдів, що являють собою розширені капіляри та венули. Цим процесам належить велика роль процесах обміну між материнським організмом і зародком.
Ранній ембріогенез
Після імплантації відбувається швидке розвиток як зародка, і його оболонок. На трофобласті утворюються ворсинки, які на ранніх стадіях онтогенезу покривають рівномірно всю поверхню бластоцисти. Первинні ворсини є безсудинними утвореннями. Поступово зовнішній шар трофобласта втрачає клітинні межі, перетворюючись на синцитії (симпласт). Внутрішній шар трофобласта зберігає клітинну будову та називається цитотрофобластом. Докладніше подальші процеси диференціації трофобласта описані у наступному розділі (див. розділ 3.2).
Одночасно з трофобластом розвивається і ембріобласт. Цей процес особливо активізується після імплантації. Клітини ембріобласту диференціюються у два вузлики: ектобластичний та ендобластичний. Незабаром у центральних частинах цих вузликів утворюються порожнини, в результаті чого ектобластичний вузлик перетворюється на ектобластічну бульбашку, а ен-
На ранніх стадіях розвитку відбувається і диференціювання мезенхіми. З одного боку клітини мезенхіми відтісняються на периферію – до трофобласту. З іншого боку відбувається скупчення мезенхімальних клітин навколо амніотичного та жовткового бульбашок, а також біля зародка. Внаслідок цього стінки бульбашок стають двошаровими, а сам зародок диференціюється в екто-, ендо-і мезодерму. Ці три зародкових листка служать вихідним матеріалом на формування надалі всіх органів прокуратури та тканин ембріона і плода.
У міру подальшого внутрішньоутробного розвитку відбувається швидке збільшення розмірів амніотичної бульбашки, яка перетворюється на амніотичну порожнину. У порожнині спостерігається швидке накопичення прозорої рідини (амніотична рідина), при цьому стінка амніотичного пляшечки наближається до ворсинчастої оболонки і, нарешті, зливається з нею. Зародок починає повертатися в порожнину амніону, а жовтковий пухирець атрофується.
Одночасно з розвитком плодових оболонок з каудального відділу первинної кишки зародка утворюється випинання - алантоїс. По ал-лантоису судини зародка "підростають" до ворсинчастої оболонки, вростаючи потім у кожну ворсину. В результаті цього безсудинний хоріон васкуля-ризується.
Після завершення початкових етапів онтогенезу ембріон оточений амніотичною рідиною та трьома оболонками: децидуальною, ворсинчастою та водною.
Децидуальна оболонка є трансформованим у зв'язку з вагітністю функціональним шаром ендометрію. На момент імплантації ендометрій знаходиться в секреторній фазі. Він складається з двох шарів: компактного та спонгіозного. Компактний шар, звернений у порожнину матки, містить вивідні протоки маткових залоз та клітини строми ендометрію, що перетворилися під час вагітності на децидуальні. Спон-гіозний шар в основному складається із залоз.
У процесі зростання плодового яйця decidua parietalis і decidua capsularis розтягуються, стоншуються та наближаються один до одного. На 4-5-му місяці вагітності плодове яйце займає вже всю порожнину матки, і decidua parietalis зливається з decidua capsularis. Навпаки, decidua basalis значно гіпертрофується та перетворюється на материнську частину плаценти. У цій частині плаценти знаходяться численні ворсини хоріону, навколо яких формується міжворсинчастий простір.
Децидуальна оболонка є для плода живильним та захисним шаром: трофічна функція її в основному здійснюється на ранніх етапах внутрішньоутробного розвитку, захисна роль найповніше виявляється високою фагоцитарною активністю. Децидуальна тканина лізує
мікроорганізми та інактивує їх токсини, бере також участь у синтезі вуглеводів, ліпідів та білків. У ній відбувається синтез пролактину та простагландинів. Таким чином, децидуальній оболонці належить дуже важлива роль в імплантації та подальшому розвитку ембріона та плода.
Амніон (водна оболонка) звернена до плода. Вона вистилає плаценту і переходить на пуповину, зливаючись в ділянці пупкового кільця зі шкірою плода. Макроскопічно амніон є тонкою напівпрозорою мембраною. У процесі ембріогенезу амніон розвивається з ектобластичної бульбашки. З ектодерми формується епітелій амніону, з мезодерми – сполучнотканинна основа.
На ранніх стадіях розвитку епітелій амніону представлений великими полігональними клітинами, з 3-го місяця вагітності стає кубічним. Епітеліальні клітини амніону містять ліпіди, полісахариди,
протеїни, фосфорні сполуки, а також ряд ферментів, що беруть участь у процесах метаболізму та обміні стероїдних гормонів.
Амніон разом із гладким хоріоном бере активну участь в обміні навколоплідних вод, а також у параплацентарному обміні. За своїми фізичними властивостями плодові оболонки відрізняються одна від одної. Так як амніотична оболонка дуже щільна і витримує тиск у кілька разів більше, ніж гладкий хоріон, під час пологів розрив гладкого хоріону настає раніше, ніж амніону.
ПЛАЦЕНТУ
Плаценталюдину має гемохоріальний тип будови. Цей тип плацентації характеризується наявністю безпосереднього контакту материнської крові з хоріоном внаслідок порушення цілісності децидуальної оболонки матки з розтином її судин.
Виникнення гемохоріальної плаценти в еволюції є найвищою стадією. У ній відображені найскладніші взаємини функціональних систем матері та плоду.
Розвиток та функції плаценти
Основною частиною плаценти є ворсини хоріону – похідні тро-фобласту. На ранніх етапах онтогенезу трофобласт утворює протоплазматичні вирости, що складаються з клітин цитотрофобласта (первинні ворсини). Первинні ворсини не мають судин, і надходження поживних речовин і кисню до організму зародка з материнської крові, що оточує їх, відбувається за законами щодо простих законів осмосу і дифузії. До кінця 2-го тижня вагітності у первинні ворсини вростає сполучна тканина та утворюються вторинні ворсини. Їхню основу становить сполучна тканина, а зовнішній покрив представлений епітелієм (трофобласт). Як первинні, і вторинні ворсини рівномірно розподіляються по поверхні плодового яйця.
Епітелій вторинних ворсин складається з двох шарів: шару цитотрофобласта (шар Лангханса) та синцитію (симпласту). Шар цитотрофобласта складається з клітин округлої форми зі світлою цитоплазмою. Ядра клітин великі. У синцитії межі клітин практично невиразні, цитоплазма темна, зерниста, із щітковою облямівкою. Ядра щодо невеликих розмірів, кулястої чи овальної форми.
З 3-го тижня розвитку зародка починається дуже важливий процес розвитку плаценти, який полягає у васкуляризації ворсин і перетворенні їх на третинні судини, що містять. Формування судин плаценти відбувається як з ангіобластів зародка, так і з пупкових судин, що ростуть з алантоїсу.
Судини алантоїсу вростають у вторинні ворсини, внаслідок чого кожна вторинна ворсина отримує васкуляризацію. Так здійснюється найважливіший процес внутрішньоутробного розвитку – васкуляризація хоріону. Встановлення алантоїдного кровообігу забезпечує інтенсивний обмін між організмами плода та матері.
▲ Перетворення вторинних ворсин у третинні розглядається як найважливіший критичний період ембріонального розвитку, оскільки алантоїс має дуже високу чутливість до дії факторів, що пошкоджують довкілля. Його пошкодження супроводжується загибеллю судин, внаслідок чого припиняється найважливіший процес- васкуляризація хоріона, а це у свою чергу призводить до загибелі зародка на ранніх термінах.
На ранніх стадіях внутрішньоутробного розвитку ворсини хоріону поступово покривають всю поверхню плодового яйця. Однак, починаючи з 2-го місяця онтогенезу на більшій поверхні плодового яйця, ворсини атрофуються, в той же час пишно розвиваються ворсини, звернені до базальної частини децидуальної оболонки. Так формуються гладкий і гіллястий хоріон.
Подальший розвиток та диференціювання хоріону характеризуються такими основними моментами. При терміні вагітності 5-6 тижнів товщина синцитіотрофобласта перевищує товщину шару Лангханса, а починаючи з терміну 9-10 тижнів синцитіотрофобласт поступово стоншується і кількість ядер у ньому збільшується. На вільній поверхні синцитіотрофобласта, зверненої до міжворсинчастого простору, стають добре помітними довгі тонкі цитоплазматичні вирости (мікроворсини), які значно збільшують резорбційну поверхню плаценти. На початку II триместру вагітності відбувається інтенсивне перетворення цитотрофобласта на синцитії, внаслідок чого на багатьох ділянках шар Лангханса повністю зникає.
Наприкінці вагітності у плаценті починаються інволюційно-дистрофічні процеси, які іноді називають старінням плаценти. З крові, що циркулює у міжворсинчастому просторі, починає випадати фібрин (фібриноїд), який відкладається переважно на поверхні ворсин. Випадання цієї речовини сприяє процесам мікро-тромбоутворення та загибелі окремих ділянок епітеліального покриву ворсин. Ворсини, покриті фібриноїдом, значною мірою виключаються з активного обміну між організмами матері та плода.
Відбувається виражене стоншення плацентарної мембрани з 25 мкм на початку вагітності до 5 мкм наприкінці її. Строма ворсин стає більш волокнистою та гомогенною. Спостерігається деяке потовщення ендотелію капілярів. У ділянках дистрофії нерідко відкладаються солі вапна. Всі ці зміни відбиваються на основних функціях плаценти (дихальної, трофічної, обмінної, ендокринної та ін.).
Поряд із процесами інволюції у плаценті протягом вагітності спостерігаються і явища протилежної властивості. Відбувається збільшення молодих ворсин, які значною мірою компенсують функцію втрачених. Проте розвиток молодих ворсин лише частково покращує функцію плаценти загалом. Внаслідок цього наприкінці вагітності спостерігається зниження функції плаценти.
Будова зрілої плаценти (рис. 3.8). Макроскопічно зріла плацента дуже нагадує товстий м'який коржик. Маса плаценти становить 500-600 г, діаметр 15-18 см, товщина 2-3 см. Плацента має дві поверхні: материнську, звернену до стінки матки, та плодову - у бік плода.

Рис. 3.8. Зовнішній вигляд зрілої плаценти, а – материнська частина; б – плодова частина.
Материнська поверхняплаценти має сірувато-червоний колір і є залишками базальної частини децидуальної оболонки.
Плодова поверхнязверху покрита блискучою амніотичною оболонкою, під якою до хоріону підходять судини, що йдуть від місця прикріплення пуповини до периферії плаценти. Основна частина плодової плаценти представлена численними ворсинами хоріону, які об'єднуються в дольчасті утворення - котиледони, або часточки. Їхнє число досягає 15-20. Частки плаценти утворюються в результаті поділу ворсин хоріону перегородками (септами), що виходять із базальної платівки. До кожної з таких часточок підходить свій великий посуд.
Мікроскопічна будова зрілої ворсини. Прийнято розрізняти два види ворсин: вільні та закріплювальні (якорні). Вільні ворсини, а таких більшість, занурені у міжворсинчастий простір децидуальної оболонки та "плавають" у материнській крові. На противагу їм якірні ворсини прикріплені до базальної децидуальної оболонки та забезпечують фіксацію плаценти до стінки матки. У третьому періоді пологів зв'язок таких ворсин із децидуальною оболонкою порушується і під впливом маткових скорочень плацента відокремлюється від стінки матки.
При мікроскопічному вивченні будови зрілої ворсини вдається диференціювати такі утворення (рис. 3.9):
Синцитій, який не має чітких клітинних кордонів;
Шар (або залишки) цитотрофобласту;
Строму ворсини;
Ендотелій капіляра, у просвіті якого добре помітні елементи
крові плода.
Під час електронної мікроскопії ворсин хоріону було встановлено, що синцитій має на своїй поверхні численні мікроворсини, які значно збільшують обмінну поверхню плаценти.
Маточно-плацентарний кровообіг. За наявності пла-
Кровотік у матці здійснюється за допомогою 150-200 материнських спіральних артерій, які відкриваються у широке міжворсинчасте простір. Спіральні артерії мають своєрідну будову, їх стінки позбавлені м'язового шару, а гирла не здатні скорочуватися та розширюватися.
Спіральні артерії мають низький судинний опір току крові. На противагу матковим артеріям, у яких виражене зниження судинного опору спостерігається з 12-13 тижнів вагітності, у спіральних артеріях, як це було встановлено за допомогою доплерометрії, цей процес має місце вже з 6 тижнів вагітності. Найбільш виражене зниження судинного опору в спіральних артеріях спостерігається в 13-14 тижнів вагітності, що морфологічно відображає завершення процесу інвазії ворсин трофоблас-ту в децидуальну оболонку.
Описані особливості гемодинаміки мають дуже велике значення у здійсненні безперебійного транспорту артеріальної крові від матері до плоду. Артеріальна кров, що вилилася, омиває ворсини хоріону, віддаючи при цьому в кров плоду кисень, необхідні поживні речовини, багато гормонів, вітаміни, електроліти та інші хімічні речовини, а також мікроелементи, необхідні плоду для його правильного росту і розвитку. Кров, що містить СО 2 та інші продукти метаболізму плода, виливається у венозні отвори материнських вен, загальна кількість яких перевищує 180.
Кровотік у міжворсинчастому просторі наприкінці вагітності досить інтенсивний і в середньому становить 500-700 мл крові на хвилину.
Особливості кровообігу в системі мати-плацента-плід.Артеріальні судини плаценти після відходження від пуповини діляться радіально відповідно до числа часток плаценти (котиледонів). Внаслідок подальшого розгалуження артеріальних судин у кінцевих ворсинах утворюється мережа капілярів, кров з яких збирається у венозну систему. Відня, в яких тече артеріальна кров, збираються у великі венозні стовбури і нарешті впадають у вену пуповини (рис. 3.10).
 |
Рис. 3.10. Кровообіг у системі мати – плацента – плід.
1 – міометрій; 2 – вільні ворсини; 3 - якірна ворсина; 4 – децидуальна оболонка; 5 - міжворсинчастий простір; 6 – спіральні артерії; 7 - хоріальна пластинка; 8 – хоріальний епітелій; 9 – амніотичний епітелій; 10 -пуповина; 11 - вена пуповини; 12 – артерії пуповини; 13 - відкладення фібриноїду.
Кровообіг у плаценті підтримується серцевими скороченнями матері та плода. Важлива роль стабільності цього кровообігу також належить механізмам саморегуляції матково-плацентарного кровообігу.
Основні функції плаценти. Платин
ембріологія.
ЕМБРІОЛОГІЯ №1
Поняття прогенезу та ембріогенезу. Періоди та основні стадії ембріогенезу у людини. Статеві клітини людини, їх структурно-генетична характеристика.
ЕМБРІОГЕНЕЗ –
ПРОГЕНЕЗ –період розвитку та дозрівання статевих клітин – яйцеклітин та сперматозоїдів, в результаті в зрілих статевих клітинах виникає гаплоїдний набір хромосом, формуються структури, що забезпечують їхню здатність до запліднення та розвитку нового організму.
ПІДЛОВІ КЛІТИНИ – ОЗНАКИ:
Гаплоїдний набір хромосом
Змінена ядерно-цитоплазматичне відношення – відношення обсягу ядра та цитоплазма
Змінено метаболізм клітини
Клітини високо диференційовані (не здатні ділитися)
ЯЙЦЕКЛІТКА (відкрита Бером ) - Має оолемму, ооплазму (цитоплазму), ядро; органоїди розвинені все крім центріолей, з включень переважає жовток.
ФІЗІОЛОГІЧНІ ОСОБЛИВОСТІ: ядро неактивне ні щодо транскрипції, ні щодо реплікації, тобто. веде себе пасивно; яйцеклітина накопичує ферменти, фактори та глікоген.
СТРУКТУРНІ ОСОБЛИВОСТІ: має кортикальний шар цитоплазми– периферична гіалоплазма з кортикальними гранулами (мукополісахариди, білки, ферменти); полярна– виділяють два полюси: анімальний (зосереджені органоїди) та вегітативний (містить< или >кількість білка)
У людини маложовткова, вториннооліголецитальна, ізолецитальна
Яйцеклітину оточують прозора (лецитальна) оболонка - zona pellucida (її утворюють білки Zp). Серед них є Zp2 білок – перешкоджає поліспермії, Zp3 рецептор до сперматозоїду.
Яйцеклітина оточена фолікулярними клітинами, які доставляють до неї харчування – формують променистий вінець.
СПЕРМАТОЗОЇД -виділяють чотири відділи: головка (містить велике ядро та акросому – видозмінена лізосома), шийка (проксимальна центріоль), тіло (мітохондріальні спіралі та дистальна центріоль), хвостик (представлений джгутиком)
Максимальна здатність до запліднення до двох діб.
Спрямована міграція сперматозоїдів визначається хемотаксисом та реотаксисом, важливими показниками при цьому є рН та слиз. Відбувається капоцитація - під дією секретів жіночих статевих шляхів сперматозоїд набувають запліднюючі здібності.
Просування сперматозоїда полегшують простогландини (діють на оболонку маткових труб)
ЕМРІОЛОГІЯ №2
Основні стадії ембріогенезу. Концепція запліднення. Характеристика запліднення людини: морфологія, необхідні умови. Концепція зиготи.
ЕМБРІОГЕНЕЗ –ембріональний розвиток людини Триває 280 днів, ділиться на три періоди: початковий (перший тиждень розвитку), зародковий (2-8 тиждень розвитку – закладення основних органів), плодовий (9 тиждень – до народження).
Ранній ембріогенез ділиться на стадії:
ЗИГОТА – початок синтезу ДНК та білка
ДРОБЛЕННЯ - початок синтезу основних типів РНК
МОРУЛА – клітини зародка тотипотентні (взаємозамінні)
БЛАСТОЦИСТА – відбувається втрата тотипотентності та клітини детермінуються до утворення зародкових та позазародкових структур.
ГАСТРУЛА – з'являються зародкові листки та стовбурові клітини
ОРГАНОГЕНЕЗ – з тканини формуються органи, йде формування зачатків органів із клітинних клонів
Оплодотворення -злиття чоловічої та жіночої статевих клітин, внаслідок чого відновлюється диплоїдний набір хромосом, характерний для даного виду і виникає якісно нова клітина – зигота (запліднена яйцеклітина або одноклітинний зародок)
Три стадії:
ПОЗИТИВНИЙ РЕОТАКСИС – рух сперматозоїда проти струму рідини, що виділяється матковими трубами
ХЕМОТКСІС – статеві клітини виділяють гормони (гамони); ♀ – гіногамони, у ♂ – андрогамони.
КАПОЦИТАЦІЯ – набуття сперматозоїдом запліднюючої здатності під дією секрету маткових труб
АКТИВАЦІЯ ЯЙЦЕКЛІТКИ – ділянка мембрани яйцеклітини, отримана від сперматозоїда проникним для іонів Na→змінюється потенціа клітини→іони Ca виходять з клітинного депо в цитоплазму→екзоцитоз кортикальних гранул. Це призводить до утворення оболонки запліднення – зміна властивостей блискучої оболонки, що перешкоджає поліспермії.
СПЕРМАТОЗОЇД ВСЕРЕДИНІ ЯЙЦЯ – триває 12 годин, у результаті утворюється зигота. У цей момент ядра статевих клітин називають пронуклеус.Їхній ядерний матрикс розпушується, оболонки зникають – стадія синкаріону. Пронуклеоси зближуються, в кожному з них відбувається подвоєння ДНК та утворення хромосом, які перемішуються та вишиковуються в метафазну пластинку першого поділу мейозу.
ДИСТАНТНА ВЗАЄМОДІЯ
КОНТАКТНА ВЗАЄМОДІЯ - сперматозоїд розсуває променистий вінець і утворює канал у блискучій оболонці. Сперматозоїд контактує з рецептором (Zp3 білок) та починається акросомна реакція – екзоцитоз вмісту акросоми для локального руйнування прозорої оболонки. Усередині головки відбувається: всередину іони Ca та Na, назовні H2. Результат – збільшення концентрації Ca запускає низку процесів, що ведуть до збільшення внутрішньоклітинного pH, а це запускає акросомну реакцію. Результат акросомної реакції - утворення каналу в прозорій оболонці, через який проходить сперматозоїд. В оолемму вбудовується мембрана сперматозоїда.
ТРЕТЯ СТАДІЯ
ЗНАЧЕННЯ СПЕРМАТОЗОЇДУ -½ хромосом у зиготі батьківські; мітохондріальний геном батька; вносить сигнальний білок дроблення; знімається блок мейозу; визначається генетичний організм.
Наслідком запліднення є зміна об'єму зиготи, деполяризація плазматичної мембрани та утворення оболонки запліднення.
ЕМБРІОЛОГІЯ №3
Поняття дроблення зародка. Характеристика дроблення людини: типи дроблення, час ембріогенезу, тривалість, умови. Будова зародка на стадії імплантації в людини.
ДРОБЛЕННЯ –послідовний мітотичний поділ зиготи на клітини (бластомери) без зростання дочірніх клітин до розміру материнської.
Клітки, що розділилися, не ростуть, не розходяться, зберігають диплоїдність. Скорочується х мітотичний цикл за рахунок G1 та G2 періодів.
Суть дроблення: утворюється багатоклітинний організм, відновлюються ядерно-цитоплазматичні відносини
У людини дроблення: повне(матеріал зиготи дробиться повністю), асинхронне(Збільшення числа бластомерів відбувається з порушенням геометричної прогресії.
ВИДИ БЛАСТОМЕРІВ: дрібні світлі та великі темні
Через 30 годин проходить перша борозна дроблення (утворюються два бластоміри)
Через 40 годин – 4 бластоміри
Через 50-60 - утворюється морула (тутова ягода)
Дрібні світлі – по периферії, утворюють трфобласт
Великі темні – усередині, утворюють ембріобласт.
На 5 добу утворюється бластула (бластоциста). Її особливість – бластоціль.
На початку з ембріобласта утворюється зародковий вузлик, а потім зародковий щиток.
На 5 добу бластоциста потрапляє до матки.
На 6-7 добу протікає перша фаза гаструляції та відбувається імплантація.
ІМПЛАНТАЦІЯ – використання зародка в слизову оболонку матки, дві стадії:
Прилипання (адгезія) - клітини трофобласти починають активно ділитися і зливаючись утворюють симпластотрофобласт - ворсин хоріону.
ПРОНИКНЕННЯ (інвазія) – симпластотрофобласт виділяє протолітичні ферменти, які послідовно руйнують епітелій, сполучну тканину та кровоносні судини слизової оболонки матки.
Спочатку зародок живиться зруйнованими тканинами матері – гістотрофний тип харчування, а потім живиться материнською кров'ю – гематрофний тип харчування.
ЕМБРІОЛОГІЯ №4
Поняття та основні механізми гаструляції. Типи гаструляції. Морфологічна та тимчасова характеристика гаструляції у людини. Будова двотижневого зародка людини. Уявлення про критичні періоди розвитку.
ГАСТРУЛЯЦІЯ -складний процес хімічних та морфогенетичних змін, що супроводжується розмноженням, зростанням, спрямованим переміщенням та диференціюванням клітин, в результаті утворюються зародкові листки: зовнішній – ектодерма, внутрішній – ентодерма, середній – мезодерма. Це джерела зародків тканин та органів, комплексу осьових органів.
СПОСОБИ ГАСТРУЛЯЦІЇ:
ІНВАГІНАЦІЯ (вп'ячування) – дно підтягується до даху
ЕПІБОЛІЯ (обростання) – характерна для великих клітин, переповнених жовтком.
ІМІГРАЦІЯ (виселення)
Делямінація (розщеплення)
У людини протікає у дві фази:
Делямінація - зародковий щиток (ембріобласт) розщеплюється на епібласт(всі зачатки крім ентодерми) та гіпобласт(ентодерма).
ІМІГРАЦІЯ (на 14-17 добу) – рух клітинних мас від головного та хвостового кінців, потім вони йдуть паралельно, утворюючи первинну смужку. Матеріал, який дрімав у головному кінці починає рухатися і йде назустріч першому потоку. Клітинний матеріал перебудовується та утворюється первинний вузлик з ямкою в центрі. Через первинний (Гензенівський) вузлик переміщається прехордальний зачаток і йде в ентодерму (у головний кінець зародка); хордальний зачаток через первинний вузлик лягає між екто-і ентодерми, утворюючи хорду. Мезодерма двома крилами підвертається через краї первинної смужки і йде всередину, розташовуючись з боків від хорди. Ектодерма завжди переміщається пластом, мезодерма – здійснює амебоподібні рухи, ентодерма відносно нерухома.
ЧИННИКИ ВИКЛИКАЮЧІ ГАСТРУЛЯЦІЮ:
Нерівномірне зростання у різних галузях зародка
Механічний (бластоміри легко змінюють форму)
Різна інтенсивність поглинання води
Різна пластичність клітинних мембран
Здатність клітин до фагоцитозу
Міжклітинні взаємодії
ЕМБРІОЛОГІЯ №5.
Поняття диференціювання зародкових листків. Уявлення про індукцію як фактор, що викликає диференціювання. Диференціювання зародкових листків та утворення зародків тканин та органів у зародка людини.
ДИФЕРЕНЦІЮВАННЯ ПЕРВИННОЇ ЕКТОДЕРМИ:
Нервова трубка(нейроцити та нейроглія сітківки ока та органу нюху, нейроцити та нейроглія головного та спинного мозку)
Нервовий гребінь та гангліозні платівки(нейроцити та нейроглія спинальних і вегетативних гангліїв, хромафінна тканина та мозкове в-во надниркових залоз)
Плакоди(Епітеліальні елементи внутрішнього вуха)
Шкірна ектодерма(епідерміс та його похідні, епітелій рогівки ока, епітелій органів ротової порожнини та його похідні, епітелій анального відділу прямої кишки, емаль та кутикула зуба, епітеліальна вистилка піхви)
Прехордальна платівка(епітелій ротової порожнини та стравоходу, епітелій трахеї, бронхів та легень)
Зародкова ектодерма
Позародкова ектодерма(епітелій амніону та пупкового канатика)
ДИФЕРЕНЦІЮВАННЯ ПЕРВИННОЇ ЕНТОДЕРМИ
Ентодерма кишкової трубки(епітелій кишечника, шлунка, печінки, підшлункової)
Жовткова ентодерма(епітелій алантоїсу та жовткового місця)
Зародкова ентодерма
Позародкова ентодерма
ДИФЕРЕНЦІЮВАННЯ МЕЗОДЕРМИ
Соміти
Міотом(скелетна м'язова тканина)
Склерот(хрящова та кісткова тканина)
Дерматом(Сполучнотканинна основа шкіри)
Мезенхіма(екзоцеломічний епітелій, сполучна тканина жовткового мішка амніону та хоріону)
Спланхнотом
Парієтальний та вісцеральний листки
Зародкова мезодерма
Сегментна ніжка нефрогонотома(епітелій гонад та сім'явиносних шляхів та нирок)
Парамезонефральна протока(Епітеліальна вистилка піхви, матки, яйцеводів)
Спланхнотом(поперечносмугаста м'язова тканина серця, коркове в-во надниркових залоз, мезотелій)
Мезенхіма(мікроглія, гладка м'язова тканина, судини, сполучні тканини, к-ки крові та кровотворних органів)
Позародкова мезодерма
Індуктори –виникають у певних ділянках зародка, зумовлюють розвиток інших ділянок у певному напрямі.
ЕМБРІОЛОГІЯ №6.
Поняття та значення позазародкових органів. Їхня поява в еволюції. Позародкові органи в людини. Освіта, будова, значення амніону, жовткового мішка, алантоїсу. Тулубна складка, її освіта, роль.
ПОЗАРОДИШЕВІ (провізорні, тимчасові) ОРГАНИ –розвиваються в процесі ембріогенезу поза тілом зародка, виконують різноманітні функції, що забезпечують зростання та розвиток самого зародка.
Деякі з цих органів, що оточують зародок, називають зародковими оболонками. До цих органів належать: амніон, жовтковий мішок, алантоїс, хоріон, плацента
Людина ці органи закладаються рано. До 11 діб розвитку починається виселення мезодерми, що заповнює порожнину бластоцисти. З епібласту виселяється позазародкова мезодерма, яка проростає перед епібластом і утворює закладку майбутньої амніотичної бульбашки. Потім по ній проростає позазародкова ектодерма. З епібласту виселяється мезенхіма, яка проростає перед гіпобластом і утворює закладку майбутнього жовткового мішка. Пізніше по ній проростає позазародкова ентодерма та утворюється жовтковий мішок.
АЛЛАНТОЇС -розвивається на 15 добу ембріогенезу як випинання стінки кишкової трубки. Проводить кровоносні судини до ворсин хоріону і, редукуючи, увійде до складу пупкового канатика.
ХОРІОН –до кінця 2 тижня ембріогенезу трофобласт починає утворювати первинні ворсини хоріону. На початку 3 тижні до них підростає мезенхіма та утворюються вторинні ворсини хоріону. Незабаром мезенхіма диференціюється у сполучну тканину та кровоносні судини, так формуються вторинні ворсини хоріону, які сформують плодову частину плаценти. Ворсини хоріона, що прилягають до основної відпадаючої оболонки, інтенсивно розростаються і утворюється гіллястий хоріон, який з'єднуючись з основною відпадаючою оболонкою, утворює плаценту.
ЖОВТОЧНИЙ МІШОК –бере участь у кровотворенні до 7-8 тижнів розвитку. Тут утворюються первинні статеві клітини, пізніше йде його еволюція і він виявляється у складі пупкового канатика у вигляді вузької трубочки.
АМНІОН –досягає великого розвитку, створює сприятливе водне середовище для розвитку зародка
???????????????????????????????????????????????????????????????????????????????????????????????????
ЕМБРІОЛОГІЯ №7
Плацента. Її значення, поява у еволюції. Типи плаценти. Плацента людини: тип, будова, функції. Структура та значення плацентарного бар'єру.
ПЛАЦЕНТУ –ворсини хоріона, що прилягають до основної відпадаючої оболонки, інтенсивно розростаються і утворюється гіллястий хоріон, який з'єднується з основною відпадаючою оболонкою, утворюючи плаценту або дитяче місце.
Зріла плацента в діаметрі 17-20 см. Товщина 2-4 см. Вага 500-600 г.
За будовою дискоїдальна, гемохоріальна, ворсинкова.
ТИПИ ПЛАЦЕНТ:
У зв'язку з особливостями проникнення ворсин хоріону в матку поділяють:
епітеліальнохоріальна – ворсини хоріону не руйнують епітелій матки (коні, свині)
десмохоріальна – ворсини хоріону руйнують епітелій матки та контактують із сполучною тканиною (жуйні та парнокопитні)
ендотеліохоріальна – ворсини хоріону руйнують епітелій матки, сполучну тканину та контактують з ендотелієм судин (хижаки та ластонгія)
гемохоріальна – ворсини руйнують епітелій матки, сполучну тканину та стінки судин та безпосередньо контактують з материнською кров'ю (примати, людина)
За характером харчування:
Хоріон поглинає з материнського організму білки, розщеплює їх до амінокислот. Синтез ембріоспецифічних білків відбувається у печінці ембріона. Сюди відносяться епітеліохоріальні та десмохоріальні плаценти. Дитинчата після народження здатні до самостійного харчування та пересування.
Хоріон поглинає амінокислоти, синтезує ембріоспецифічні білки (всі інші плаценти) дитинчата після народження порівняно довго адаптуються.
БУДОВА ПЛАЦЕНТИ:
Плодна частина
АМНІОТИЧНА ОБОЛОЧКА – складається з одношарового призматичного епітелію та позазародкової сполучної тканини.
ХОРІАЛЬНА ПЛАСТИНКА
ХОРІАЛЬНІ ВОРСИНИ
Строму (основу) хоріальної пластини та ворсин складає пухка сполучна тканина (містить аморфну речовину, аргірофтльні волокна, безліч капілярів та клітини Кащенко-Ховбауера – примітивні макрофаги). Зверху пластини покриті трофобластним епітелієм. У першому триместрі покриті цитотрофобластом, у другому покриті цито- та симпластотрофобластом. У третьому – фібриноїдом (гомогенная оксифільна маса, яка є продуктом сироватки крові та розпаду трофобластних елементів). У ході вагітності ворсини сильно збільшуються у розмірах і зростає їхнє розгалуження. Вони збираються групами по 15-16 штук і формують котилідони (група ворсин, пов'язана за допомогою ствола ворсин із хоріальною платівкою). Виділяють якірні ворсини (дотикаються до материнської тканини) і кінцеві ворсини (дотикаються до материнської крові). Утворюється сполучна тканина: септи межі між котиледонами.
Материнська частина
ОСНОВНА ВІДПАДНА ОБОЛОНКА (базальна пластина)
З'ЄДНУВАЛЬНІ СЕПТИ
ЛАКУНИ, ЗАПОВНЕНІ МАТЕРИНСЬКОЮ КРОВЮ
У сполучнотканинній слизовій оболонці з'являються децидуальні клітини (ті ж сполучнотканинні клітини великих розмірів і багаті на глікоген. Збільшується просвіт судин і гідратація слизової оболонки, з'являється виняткова фагоцитарна активність
ФУНКЦІЇ ПЛАЦЕНТИ:
Синтез БЖУ
Іони Ca, Fe, Zn, Mg, Cu, P
Депонує та містить вітаміни груп: A, B1, B2, B6, C, D, E
Фактори згортання крові
Дихальна
Трофічна
Ендокринна- Усі гормони синтезуються симпластотрофобластом. Вони необхідні для зростання та розвитку плоду, готують організм матері до пологів та лактації.
Гормони:
хоріонічний гонадотропін (підтримка жовтого тіла, що виділяється з перших днів вагітності, діє на жовте тіло та стимулює секрецію прогестеронів та естрогенів, які діють на ендометрій, викликаючи зростання зародка
хоріонічний соматомаммотропін (Діє на молочну залозу і жовте тіло, викликаючи зміни в організмі матері)
кортиколіберін (Визначає термін настання пологів)
Імунологічний захист
МЕХАНІЗМИ ЗАХИСТУ ПЛОДУ: симпластотрофобласт- синтезує білки матері, що гальмують імунну відповідь, гормони плаценти- пригнічують материнські лімфоцити, фібриноїд плаценти
Бар'єрна (захисна)- Виражена слабо, тому проникають наркотики, алкоголь, нікотин, отрути, ліки (антибіотики, сульфаніламіди, анальгетики), віруси, бактерії
ПЛАЦЕНТАРНИЙ БАР'ЄР:бар'єр між кров'ю матері та плоду. До бар'єру підходять: цитотрофобласт з його базальною мембраною, сполучна тканина, строма ворсини, ендотелій кровоносної судини та її базальна мембрана. З материнської сторони кров надходить у міжворсинковий простір через 30 спірально звивистих артерій матки за тиском 70-80 мм рт. ст. Материнська кров омиває ворсини, а потім накопичується на дні плацентарних відсіків, звідки відноситься через крайовий синус у маткові вени. Об'єм крові 150 мл.
ЕМБРІОЛОГІЯ №8
Поняття та значення плацентації. Плацентація у людини: тимчасова та морфологічна характеристика. Тип та будова сформованої плаценти.







